FGFR-3 e o crescimento ósseo
•
0 gostou•227 visualizações

O documento discute como os estudos recentes estenderam nosso entendimento sobre os fatores que afetam o crescimento ósseo e estatura, incluindo sinalizadores secretores e receptores, bem como fatores parácrinos e autócrinos que afetam localmente as placas de crescimento. Mutações no receptor do fator de crescimento de fibroblastos 3 (FGFR-3) foram associadas a altura alterada.

Recomendados
Recomendados
Mais conteúdo relacionado
Mais procurados
Mais procurados (20)
Destaque
Semelhante a FGFR-3 e o crescimento ósseo
Semelhante a FGFR-3 e o crescimento ósseo (20)
Mais de Van Der Häägen Brazil
Mais de Van Der Häägen Brazil (20)
Último
Último (11)
FGFR-3 e o crescimento ósseo
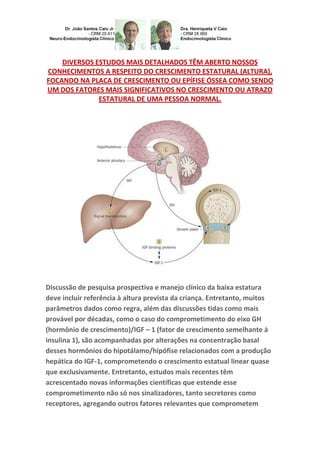
- 1. DIVERSOS ESTUDOS MAIS DETALHADOS TÊM ABERTO NOSSOS CONHECIMENTOS A RESPEITO DO CRESCIMENTO ESTATURAL (ALTURA), FOCANDO NA PLACA DE CRESCIMENTO OU EPÍFISE ÓSSEA COMO SENDO UM DOS FATORES MAIS SIGNIFICATIVOS NO CRESCIMENTO OU ATRAZO ESTATURAL DE UMA PESSOA NORMAL. Discussão de pesquisa prospectiva e manejo clínico da baixa estatura deve incluir referência à altura prevista da criança. Entretanto, muitos parâmetros dados como regra, além das discussões tidas como mais provável por décadas, como o caso do comprometimento do eixo GH (hormônio de crescimento)/IGF – 1 (fator de crescimento semelhante à insulina 1), são acompanhadas por alterações na concentração basal desses hormônios do hipotálamo/hipófise relacionados com a produção hepática do IGF-1, comprometendo o crescimento estatual linear quase que exclusivamente. Entretanto, estudos mais recentes têm acrescentado novas informações científicas que estende esse comprometimento não só nos sinalizadores, tanto secretores como receptores, agregando outros fatores relevantes que comprometem
- 2. localmente as placas de crescimento ou epífises ósseas por secreções parácrinas e autócrinas entre outras (sinalização parácrina é uma forma de comunicação célula-célula em que uma célula produz um sinal de indução e alterações em células vizinhas, que alteram o comportamento ou a diferenciação dessas células. Moléculas de sinalização conhecidas como fatores parácrinos difundem através de uma distância relativamente curta (de ação local), em oposição a endócrinos (hormônios que viajam consideravelmente mais longas distâncias, através do sistema circulatório), sinalização autócrina é uma forma de sinalização celular no qual uma célula segrega um hormônio ou mensageiro químico (o chamado agente autócrino) que se liga a receptores autócrinos na mesma célula, levando a alterações na célula). Essas possibilidades aumentam muito a abertura das portas do conhecimento cientifico, explicando situações antes questionáveis. O receptor do fator de crescimento de fibroblastos 3 (FGFR-3) é um regulador negativo da placa de crescimento na ação de condrogenesis.
- 3. Conseqüentemente, mutações ativadoras no receptor do fator de crescimento de fibroblastos 3 (FGFR-3 no alongamento vem prejudicar a composição óssea em pacientes com acondroplasia, hipocondroplasia ou displasia tanatofórica (displasia tanatofórica é um grave comprometimento esquelético onde existe desordem caracterizada por uma desproporcionalidade com pequena caixa torácica, membros extremamente curtos e dobras de pele extra sobre os braços e pernas, ou seja, os segmentos dos membros não são proporcionais entre si como em indivíduos normais). Um relatório publicado em 2015 identificaram uma mutação que ativa o receptor do fator de crescimento de fibroblasto 3 (FGFR-3) em uma família com presença autossômica com baixa estatura proporcional dominante. Por outro lado, heterozigotos e homozigotos com mutações que inativam receptor do fator de crescimento de fibroblastos 3 (FGFR-3) foram relatados em indivíduos com estatura alta. Na placa de crescimento, o FGF- fator de crescimento de fibroblastos, a sinalização através de receptor do fator de crescimento de fibroblastos 3 (FGFR-3) conduz à ativação de diversas vias, incluindo a cinase de proteína (uma cinase de proteína é uma cinase de enzima que modifica outras proteínas pela adição química de grupos de fosfato denominado (fosforilação)), ativada através de mitogênio (MAPK) e Janus transdutor de sinal-quinase e vias ativadoras de transcrição (JAK-STAT), de FGFR-3 sinalização regulando negativamente o crescimento, diminuindo a
- 4. proliferação na zona proliferativa, a diminuição da produção da matriz extracelular, acelerar o início da diferenciação hipertrófica e diminuindo o tamanho dos condrócitos hipertróficos. Esses efeitos são, pelo menos em parte, devido às interações com outros fatores parácrinos. Por tanto, a complexidade do crescimento através da estimulação por diversos hormônios e enzimas ou mesmo o bloqueio é talvez tão ou mais relevante devido sua ação na placa de crescimento, assim como o eixo GH/IGF-1. Dr. João Santos Caio Jr. Endocrinologia – Neurocientista-Endócrino CRM 20611 Dra. Henriqueta V. Caio Endocrinologista – Medicina Interna CRM 28930 COMO SABER MAIS: 1. O eixo GH/IGF-1 e sua relação com o osso que é um tecido conjuntivo dinâmico que sofre um processo contínuo de reabsorção e renovação... Outros Assuntos http://clinicascaiomutacoes.blogspot.com.br/ 2. As principais células que mediam este processo incluem as células osteoprogenitoras que contribuem para a manutenção da população de osteoblastos, os osteoblastos sintetizam a matriz óssea, osteócitos, que influenciam a estrutura do osso e a resposta à carga mecânica e os osteoclastos que promovem a reabsorção óssea... Outros Assuntos http://clinicascaiocrescereisim.blogspot.com/
- 5. 3. A maturação e função dos osteoblastos requer um espectro de proteínas de sinalização, incluindo os morfogênicos, hormonais, fatores de crescimento, citocinas, proteínas da matriz e fatores de transcrição que atuam de um modo específico-temporal (Aubin et al. 2006)... Outros Assuntos http://clinicascaioimunidade.blogspot.com.br/ AUTORIZADO O USO DOS DIREITOS AUTORAIS COM CITAÇÃO DOS AUTORES PROSPECTIVOS ET REFERÊNCIA BIBLIOGRÁFICA. Referências Bibliográficas: Caio Jr., Dr. João Santos. Endocrinologista – Neuroendocrinologista e Dra. Caio, Henriqueta V. Endocrinologista – Medicina Interna, Van Der Häägen Brasil – São Paulo – Brasil; Abe T, Nomura S, Nakagawa R, Fujimoto M, Kawase I & Naka T 2006 Osteoblast differentiation is impaired in SOCS-1-deficient mice. Journal of Bone and Mineral Metabolism24 283–290. (doi:10.1007/s00774-006-0685-0) Adams TE, Hansen JA, Starr R, Nicola NA, Hilton DJ & Billestrup N 1998 Growth hormone preferentially induces the rapid, transient expression of SOCS-3, a novel inhibitor of cytokine receptor signaling. Journal of Biological Chemistry 273 1285–1287. (doi:10.1074/jbc.273.3. 1285) Akeno N, Robins J, Zhang M, Czyzyk-Krzeska MF & Clemens TL 2002 Induction of vascular endothelial growth factor by IGF-I in osteoblast-like cells is mediated by the PI3K signaling pathway through the hypoxia- inducible factor-2alpha. Endocrinology 143 420–425. (doi:10.1210/en.143.2.420) Alexander WS, Starr R, Fenner JE, Scott CL, Handman E, Sprigg NS, Corbin JE, Cornish AL, Darwiche R, Owczarek CM et al. 1999 SOCS1 is a critical inhibitor of interferon gamma signaling and prevents the potentially fatal neonatal actions of this cytokine. Cell 98 597–608. (doi:10.1016/S0092-8674(00)80047-1) Andiran N & Yordam N 2007 TNF-alpha levels in children with growth hormone deficiency and the effect of long-term growth hormone replacement therapy. Growth Hormone & IGF Research 17 149–153. (doi:10. 1016/j.ghir.2007.01.002) Aubin JE, Lian JB & Stein GS 2006 Bone formation: maturation and functional activities of osteoblast lineage cells. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism, 6th ed., pp 20–29. Ed. Favus MJ. Washington DC: American Society for Bone and Mineral Research. Auernhammer CJ, Bousquet C & Melmed S 1999 Autoregulation of pituitary corticotroph SOCS-3 expression: characterization of the murine SOCS-3 promoter. PNAS 96 6964–6969. (doi:10.1073/pnas.96.12.6964) Baker J, Liu JP, Robertson EJ & Efstratiadis A 1993 Role of insulin-like growth factors in embryonic and postnatal growth. Cell 75 73–82. (doi:10. 1016/S0092-8674(05)80085-6) Ballesteros M, Leung KC, Ross RJ, Iismaa TP & Ho KK 2000 Distribution and abundance of messenger ribonucleic acid for growth hormone receptor isoforms in human tissues. Journal of Clinical Endocrinology and Metabolism 85 2865–2871. (doi:10.1210/jc.85.8.2865) Barnard R, Haynes KM, Werther GA & Waters MJ 1988 The ontogeny of growth hormone receptors in the rabbit tibia. Endocrinology 122 2562–2569.(doi:10.1210/ endo-122-6-2562) Beamer WH & Eicher EM 1976 Stimulation of growth in the little mouse. Journal of Endocrinology 71 37–45. (doi:10.1677/joe. 0.0710037) Behringer RR, Lewin TM, Quaife CJ, Palmiter RD, Brinster RL & D’Ercole AJ 1990 Expression of insulin- like growth factor I stimulates normal somatic growth in growth hormone-deficient transgenic mice. Endocrinology 127 1033–1040. (doi:10.1210/endo-127-3-1033). Site Van Der Häägen Brazil www.vanderhaagenbrazil.com.br
- 6. www.clinicavanderhaagen.com.br http://drcaiojr.site.med.br http://dracaio.site.med.br Joao Santos Caio Jr http://google.com/+JoaoSantosCaioJr google.com/+JoãoSantosCaioJrvdh google.com/+VANDERHAAGENBRAZILvdh Video http://youtu.be/woonaiFJQwY VAN DER HAAGEN BRAZI Instagram https://instagram.com/clinicascaio/ Wordpress http://crescimentodna.wordpress.com Google Maps: http://maps.google.com.br/maps/place?cid=5099901339000351730&q=Van+Der+Haagen+Brasil&hl=pt&sll=- 23.578256,46.645653&sspn=0.005074,0.009645&ie =UTF8&ll=-23.575591,-46.650481&spn=0,0&t = h&z=17