Baixado 409 vezes


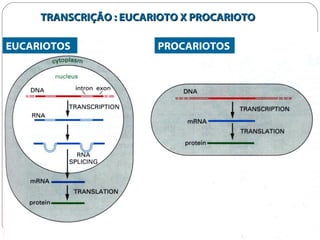
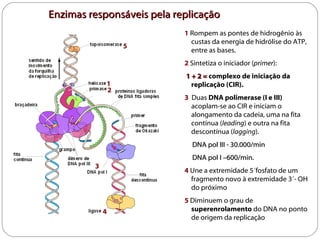
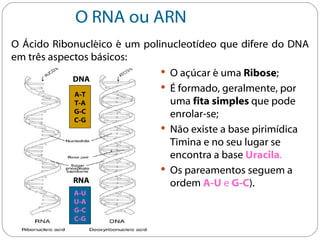






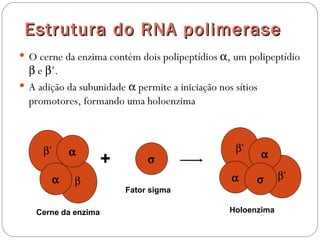
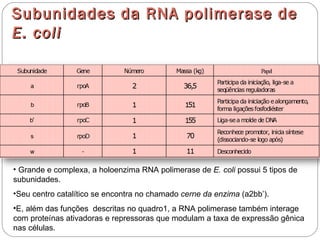
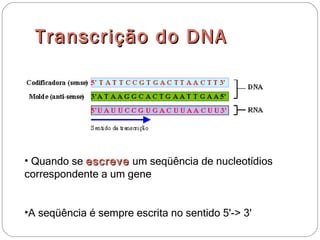
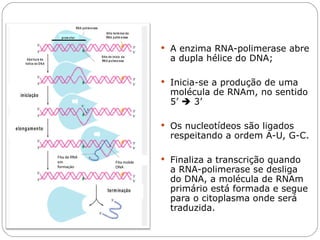


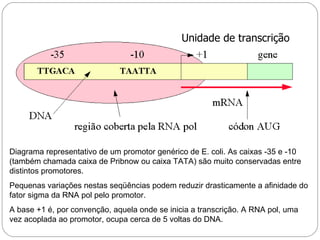


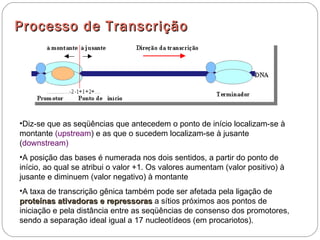
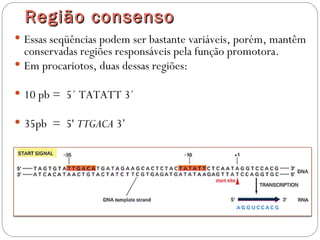





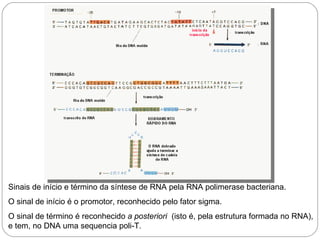


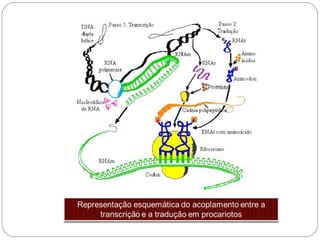



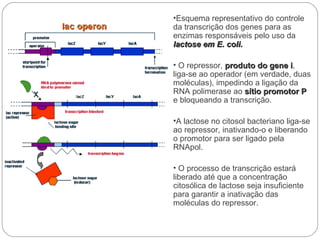


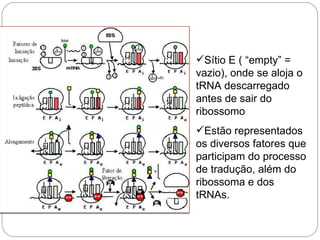
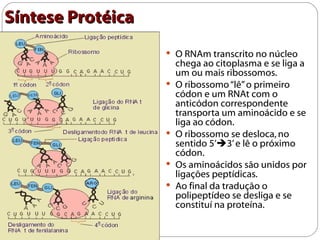
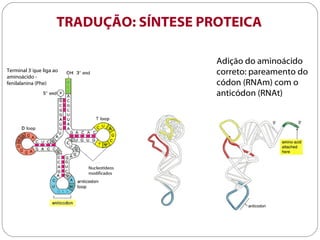
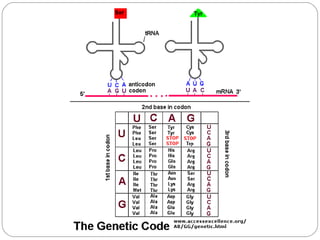

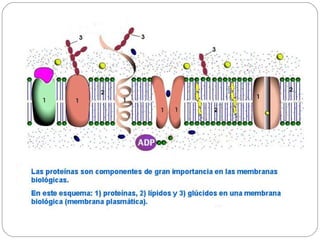

O documento descreve os processos de transcrição e regulação gênica em bactérias. A transcrição envolve as etapas de iniciação, alongamento e término, mediadas pela RNA polimerase e reconhecimento de seqüências específicas de DNA como promotores e terminadores. A expressão gênica pode ser regulada em diferentes níveis como transcrição, processamento e tradução do RNA.












![Aula Biologia: Bases da Ecologia [1° Ano do Ensino Médio]](https://cdn.slidesharecdn.com/ss_thumbnails/mdulo4-basesdaecologia-170208190312-thumbnail.jpg?width=640&height=640&fit=bounds)



































![Aula 1 - Biodiversidade e os Recursos Genéticos [Salvo automaticamente].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/aula1-biodiversidadeeosrecursosgenticossalvoautomaticamente-220804193901-27541340-thumbnail.jpg?width=640&height=640&fit=bounds)






































