Baixado 227 vezes





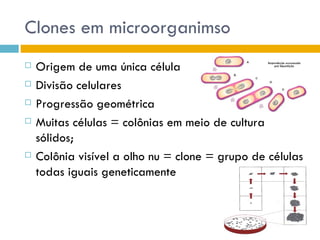





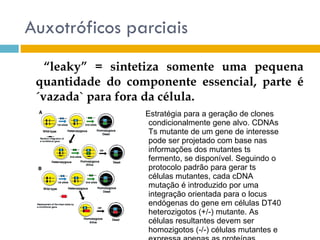


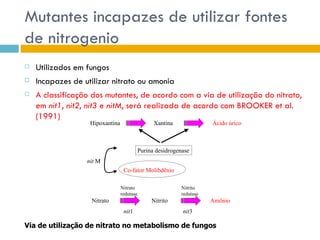
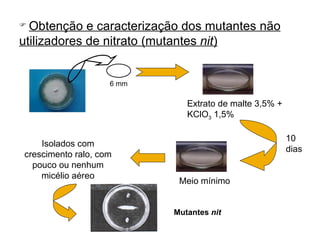
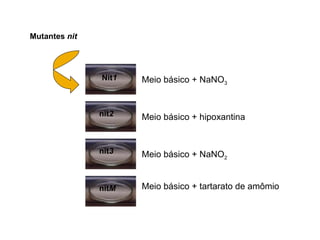




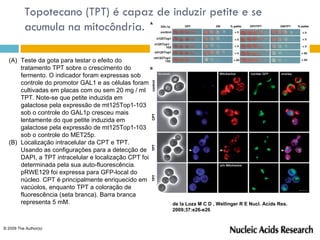
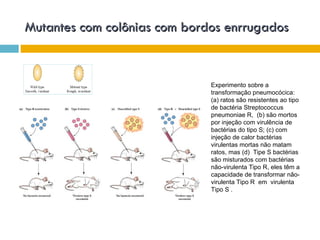
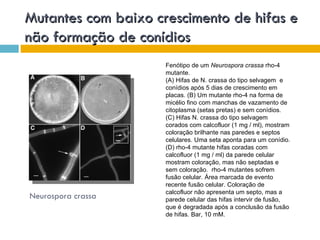

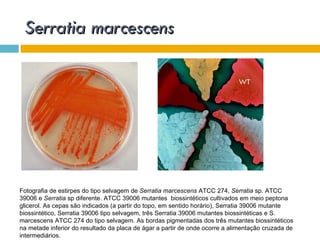
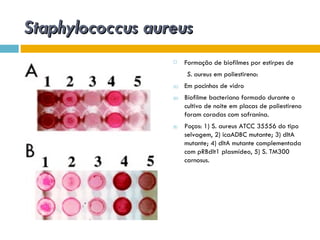



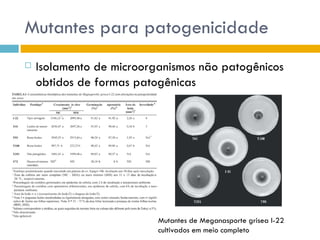




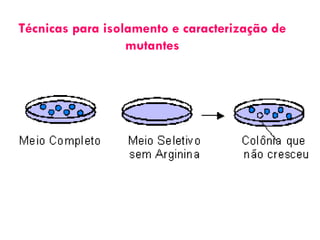

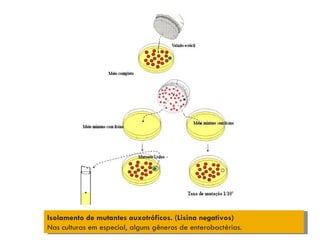

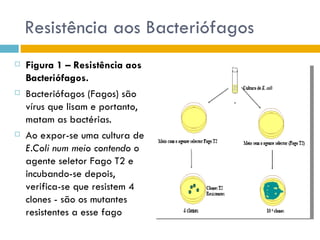
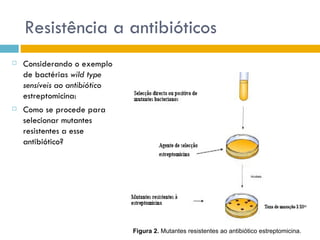


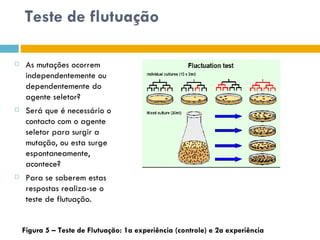
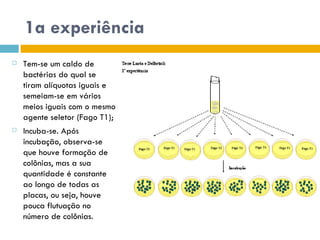
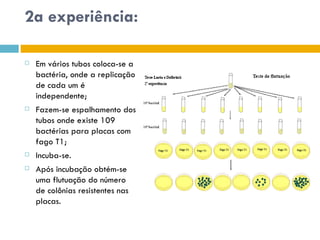


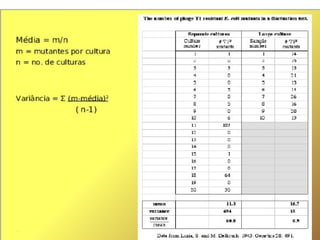

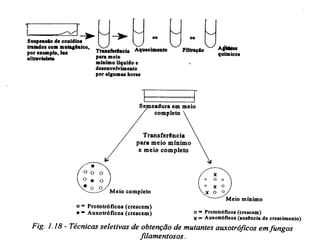
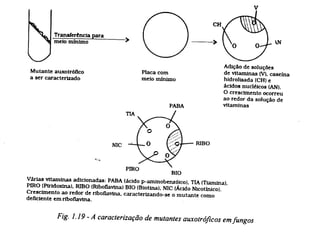
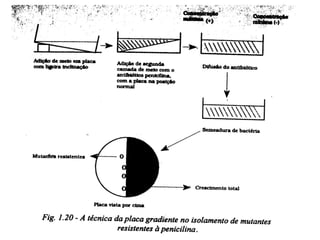


O documento discute como as mutações em microrganismos geram variabilidade e contribuem para a evolução. As principais formas de variação genética incluem mutações, recombinações e aquisição de novos genes. Os mutantes auxotróficos, resistentes a antibióticos e com alterações morfológicas são úteis para estudos genéticos de microrganismos.




































































![Aula 1 - Biodiversidade e os Recursos Genéticos [Salvo automaticamente].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/aula1-biodiversidadeeosrecursosgenticossalvoautomaticamente-220804193901-27541340-thumbnail.jpg?width=640&height=640&fit=bounds)






































