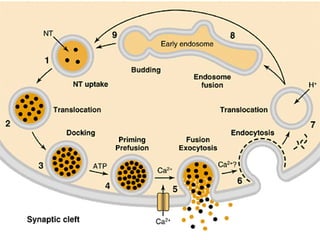
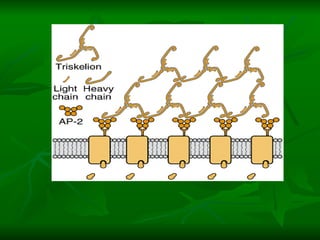
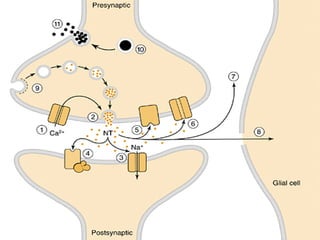
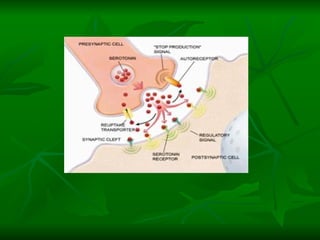
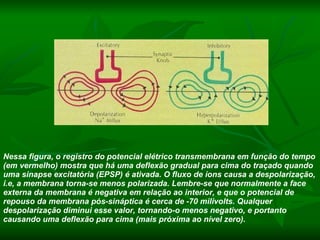
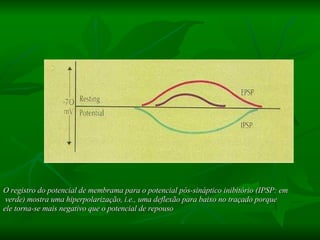
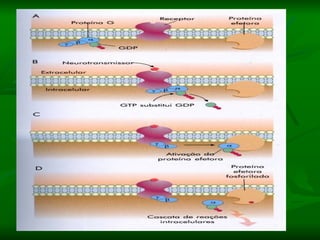
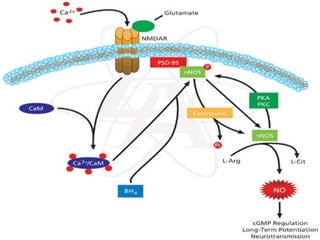
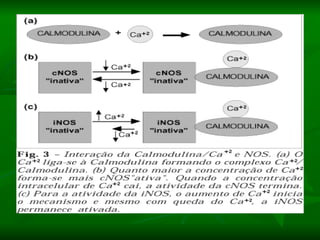
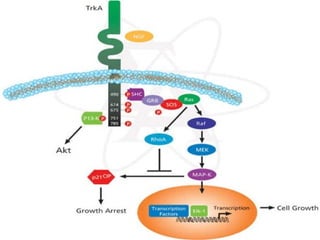
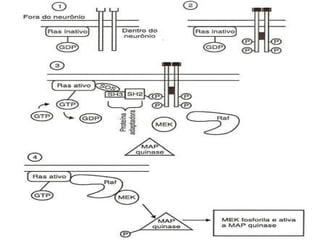
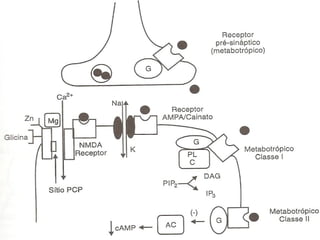
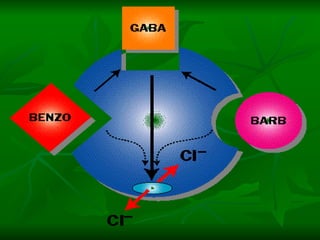
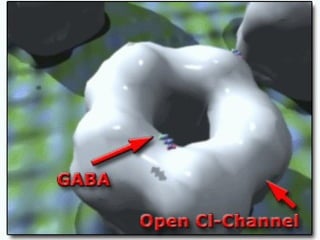
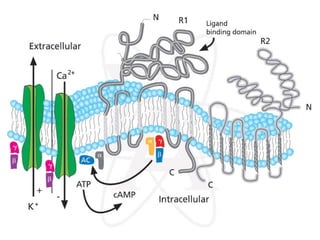
1. O documento descreve os processos de neurotransmissão, incluindo a liberação do neurotransmissor na fenda sináptica, a ligação aos receptores na membrana pós-sináptica e as respostas celulares.
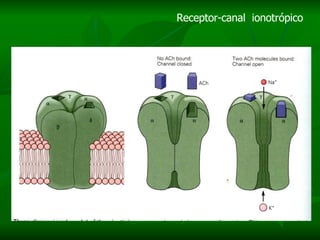
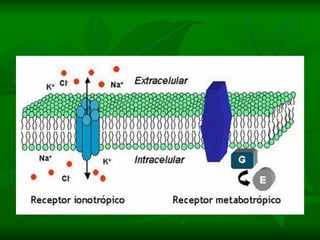
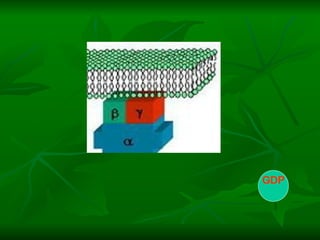
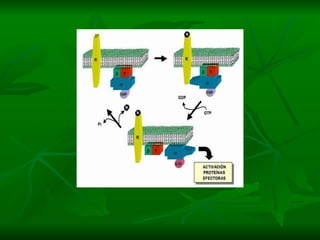
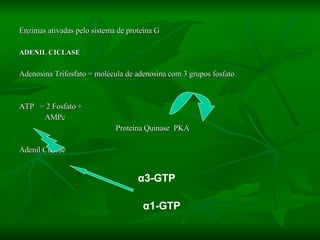
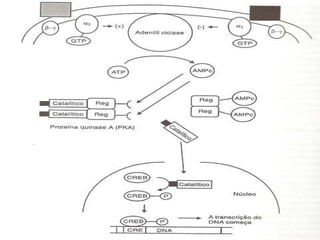
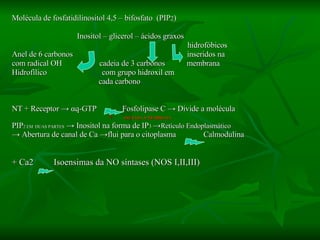
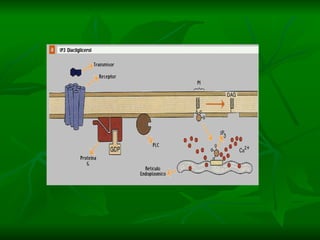
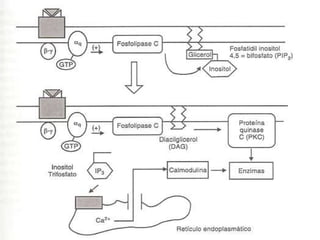
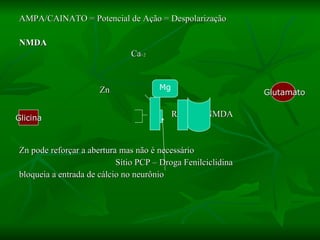
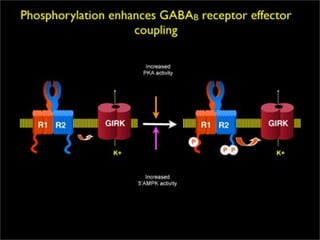
2. São descritos dois tipos principais de receptores: ionotrópicos, que abrem canais iônicos, e metabotrópicos, que ativam segundos mensageiros.
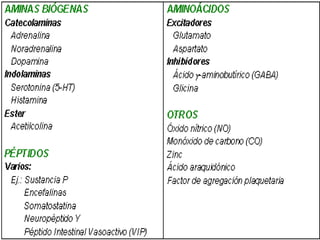
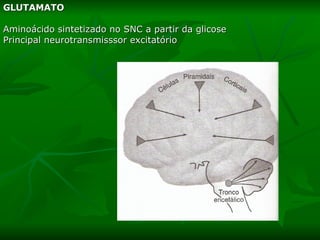
3. Os principais sistemas neurotransmissores, como o glutamato, são descritos em termos de localização, tip