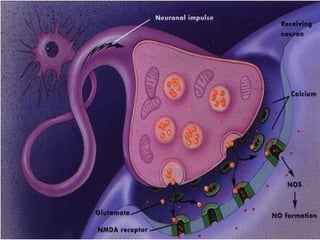
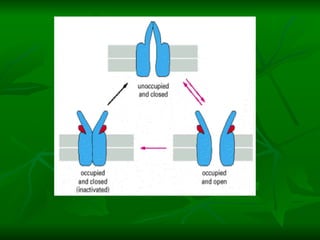
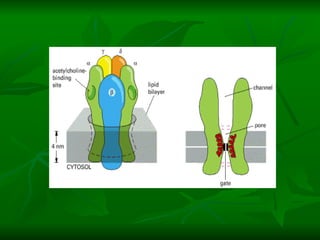
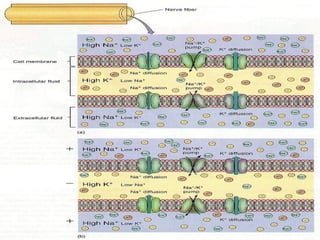
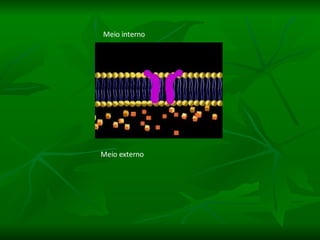
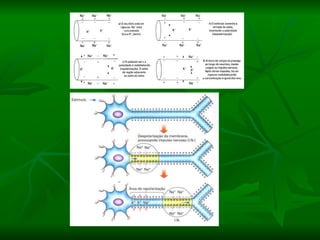
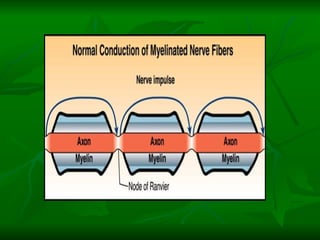
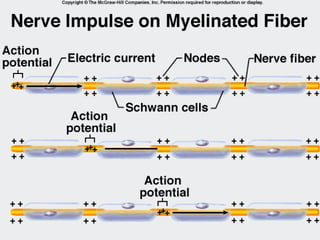
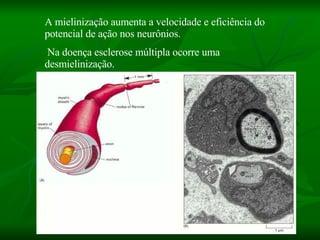
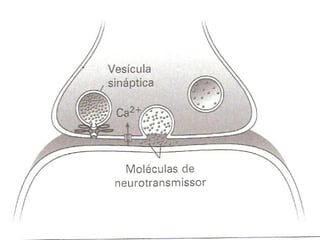
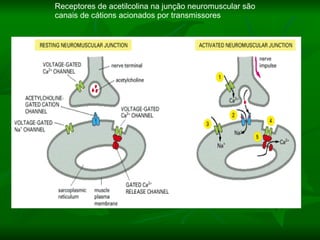
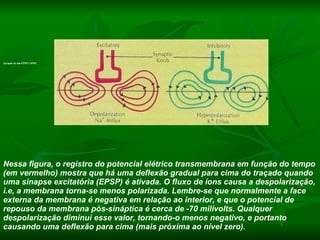
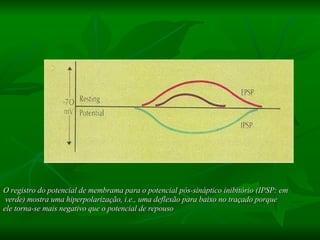
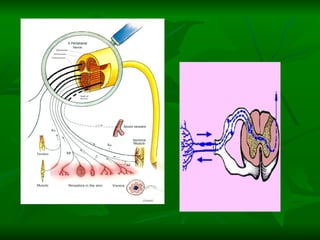
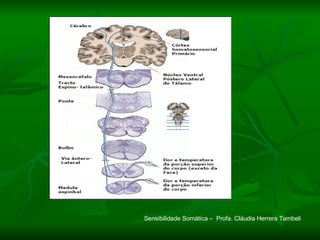
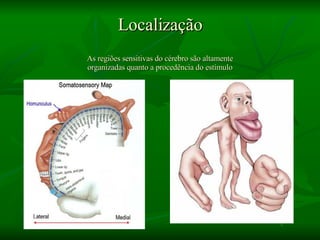
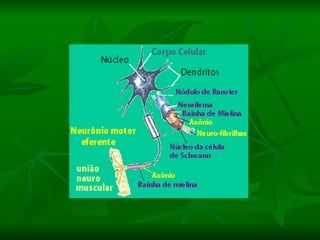
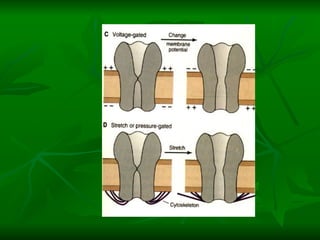
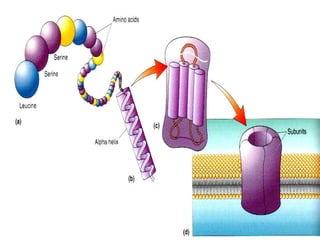
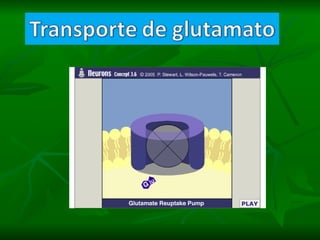
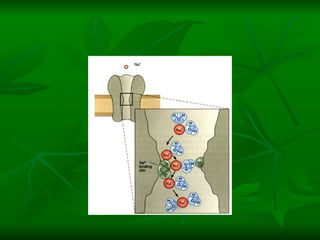
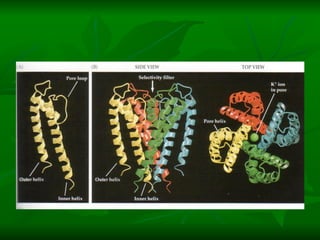
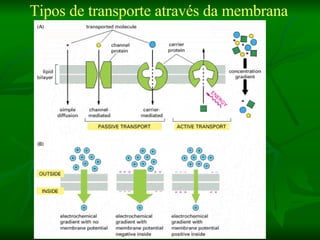
1. O documento descreve as principais características estruturais e funcionais dos neurônios e das sinapses. Apresenta a membrana celular, organelas, citoesqueleto, axônios, botões terminais e neurotransmissores.
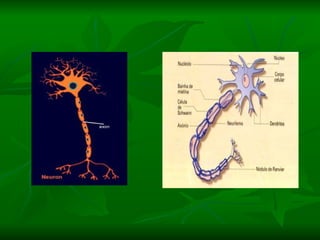
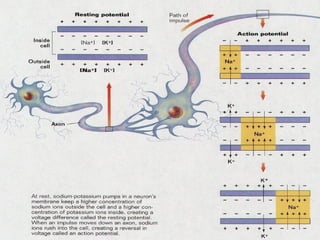
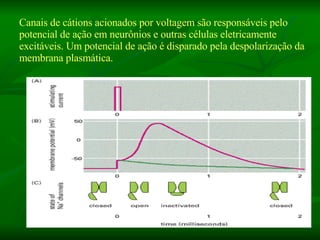
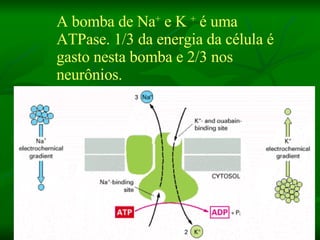
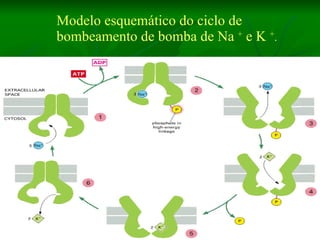
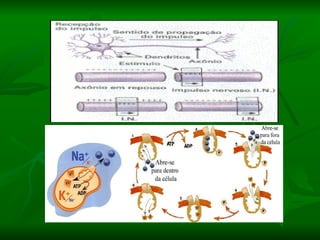
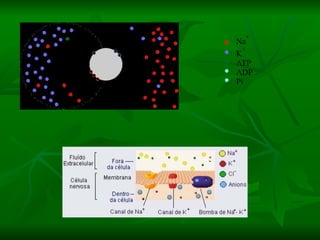
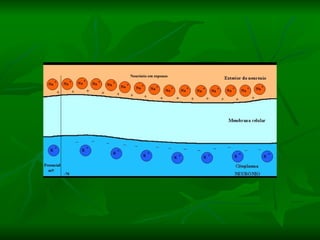
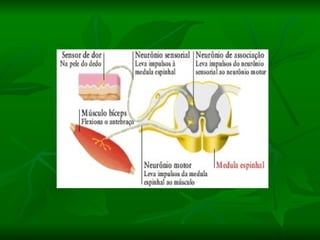
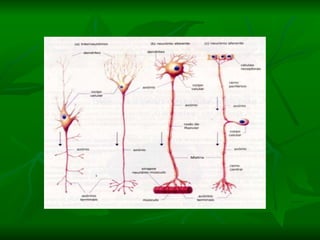
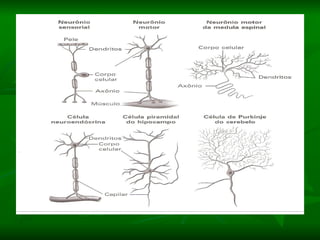
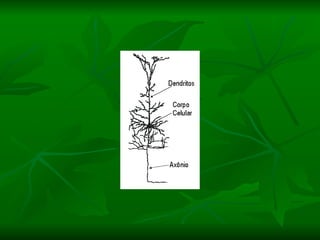
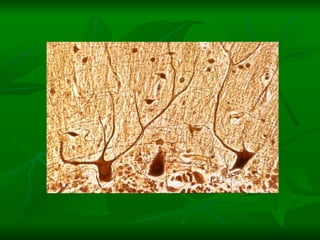
2. Os neurônios são células especializadas na comunicação e processamento de informações através do sistema nervoso. Possuem dendritos para receber estímulos, corpo celular com núcleo e organelas, e axônios para conduzir o potencial de ação













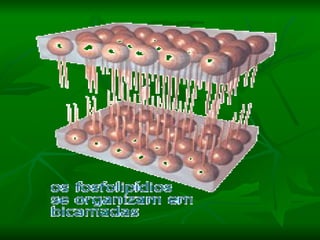
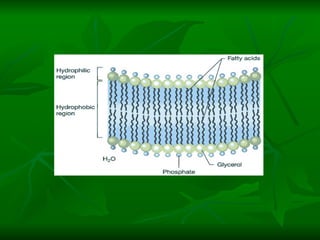
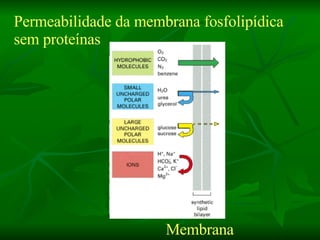

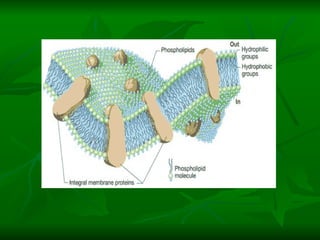
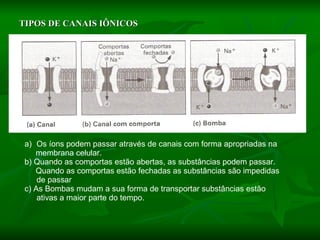
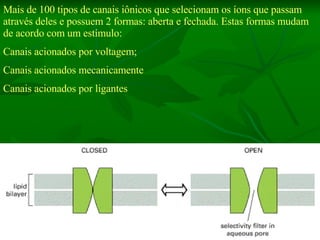
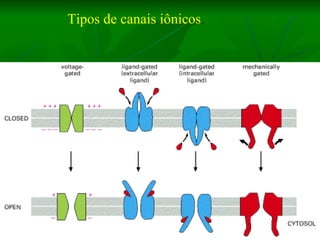
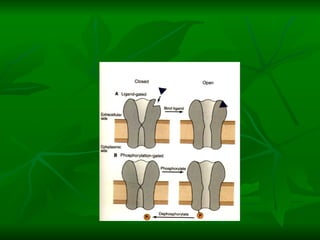


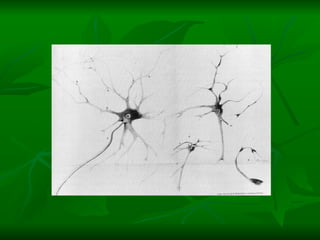
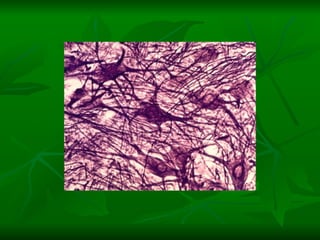
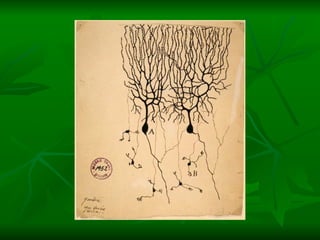

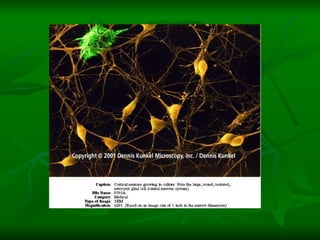

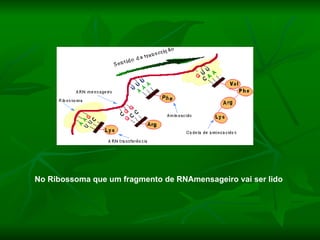
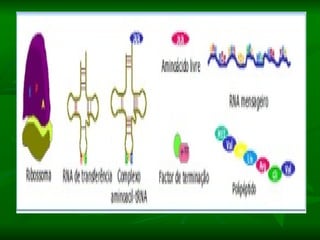
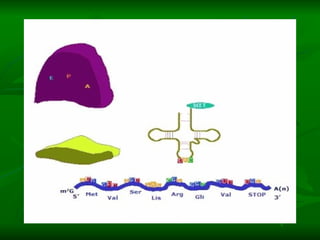
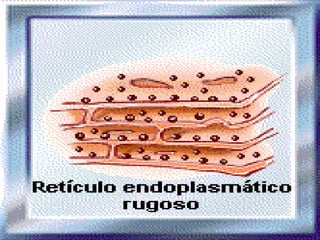

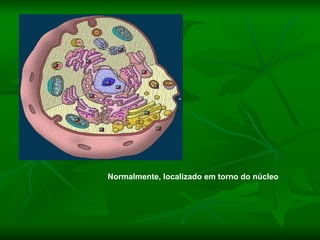
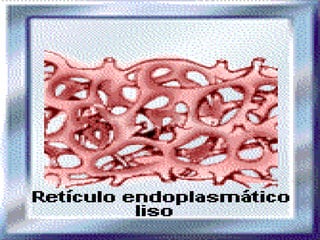

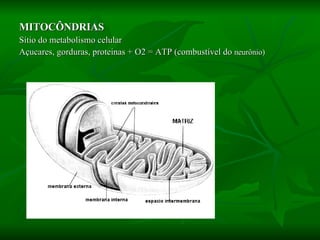
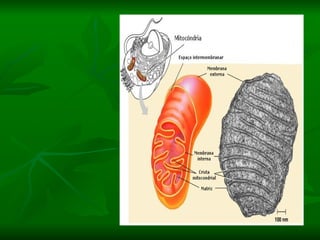

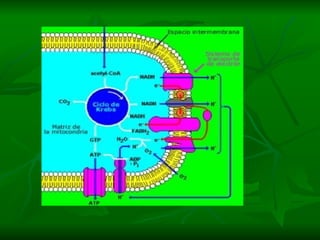
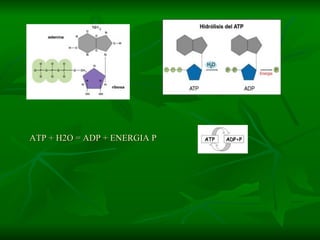


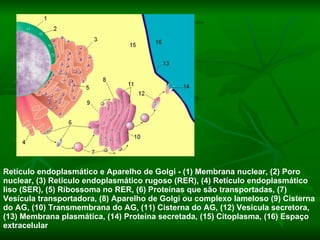



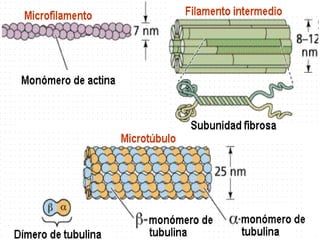

![NEUROFILAMENTOS Filas longas de apêndices protéicos, dispostos uns sobre os outros. Responsável pela a transferência de produtos neuronais dentro do citoplasma] Envolvido no controle e transporte de proteínas - “corridas curtas, não maratonas” – processo intermitente. Envolvido também no apoio estrutural Doença de Alhzheimer – neurofilamentos se dispõem como um emaranhado](https://image.slidesharecdn.com/cpia-de-neurnio-3-1217540035639182-8/85/CoPia-De-NeuroNio-3-70-320.jpg)