Baixado 278 vezes





















![Arlindo Ugulino Netto; Raquel Torres Bezerra Dantas ● MEDRESUMOS 2016 ● CEF
26
www.medresumos.com.br
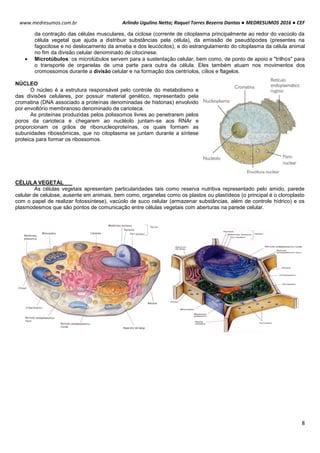
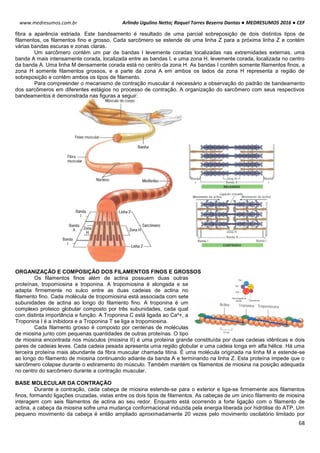
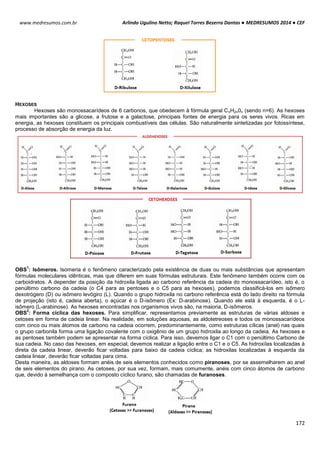
SITEMA DE CONTROLE DO CICLO CELULAR
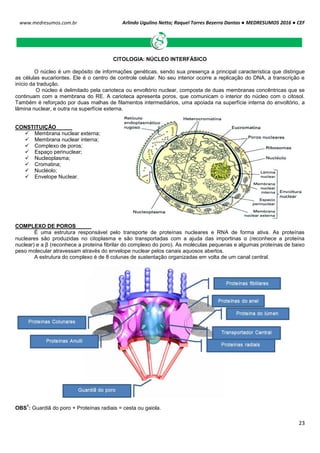
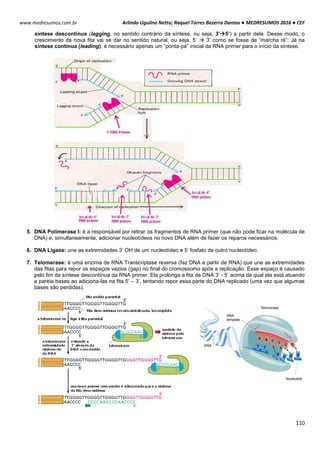
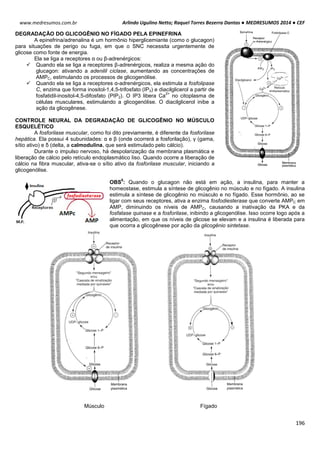
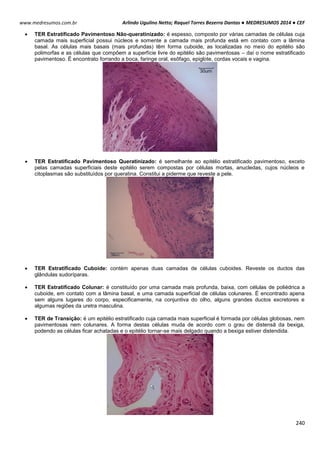
O ciclo celular é controlado e acompanhado pelas ciclinas (reguladora mitótica ou G1) e pelas proteinoquinases
[dependentes de ciclinas (CDK), com ação de proliferação]. Essas CDK (proteinoquinases dependentes de ciclinas)
controlam o sistema do processo de divisão, observando se é para a célula parar ou continuar a divisão.
As ciclinas sofrem acumulação (ativam a CDK correspondente) e degradação periódicas (via ubiquitina-
proteossoma).
A ciclina sozinha não consegue ativar a CDK, para isso tem que haver fosforilação e desfosforilação em sítios
específicos para tornarem-se enzimaticamente ativas.
A síntese de ciclinas e a presença de fatores de crescimento estão relacionadas, por isso se não houver um fator
de crescimento, haverá uma deficiência de ciclinas decorrendo em um bloqueio no ciclo celular e a célula entra em G0.
MITOSE
A mitose caracteriza-se pela condensação dos cromossomos, ocorrendo devido a fosforilação da histona 1. As
consequências observadas neste processo são:
Acúmulo de ciclinas e sua ligação para formar o fator protetor de maturação (MPF).
A condensação do material genético é realizada por condensinas, proteínas fosforiladas por CDK1. A citocinese
gerada pela desativação do MPF coordena a divisão citoplasmática e nuclear da célula.
Acumulação da ciclina Associação com a CDK
correspondente
Ativação da CDK
CDK ativa: fosforilaçãoEventos do ciclo celular](https://image.slidesharecdn.com/medresumos2016-cef-170904035900/85/Medresumos-2016-cef-26-320.jpg)

![Arlindo Ugulino Netto; Raquel Torres Bezerra Dantas ● MEDRESUMOS 2016 ● CEF
30
www.medresumos.com.br
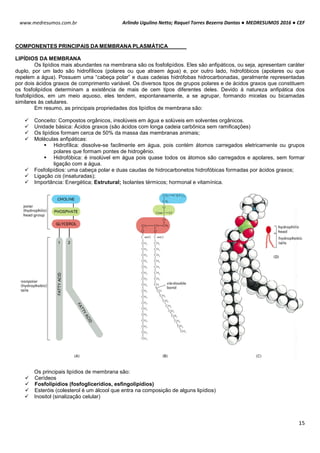
CITOLOGIA: RIBOSSOMOS
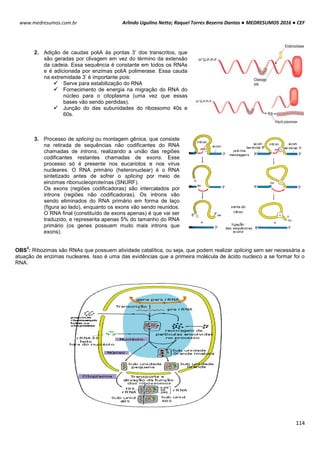
Ribossomos são organelas citoplasmáticas encontradas em procariotos e eucariotos. Eles são amplos
complexos de proteínas (proteínas ribossomais) e moléculas de RNAs ribossômicos (rRNAs), sendo 3 moléculas de
RNAr nos procariotos e quatro nos eucariotos. Esses complexos de proteínas [RNAr] são chamados subunidades e são
produzidos no nucléolo. A principal função dos ribossomos é servir de sítio de tradução, ou seja, síntese de proteínas
(reunião de aminoácidos em proteínas) uma vez que 2 subunidades (uma grande e uma pequena) são unidas pelo
mRNA em uma sequência especifica de aminoácidos ou uma cadeia polipeptídica.
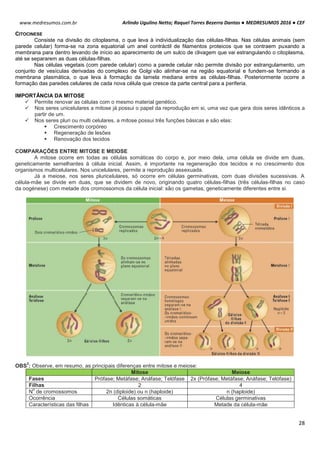
São encontrados nas células sob duas formas: livres ou associados ao
retículo endoplasmático.
Livres: encontrados no citoplasma, pode ocorrer com um único
ribossomo ou em grupos conhecidos como polissomos. Responsável
por proteínas que estão em solução no citoplasma.
Associados ao reticulo: encontrados associados à membrana exterior
do retículo endoplasmático. Responsáveis pelas proteínas que formam
membranas, que são estocadas em vesículas no citoplasma ou
exportadas para o exterior da célula.
Destino das proteínas dos ribossomos livres: núcleo, peroxissomos,
mitocôndrias e cloroplastos.
Destino das proteínas aderidas no Retículo Endoplasmático:
lisossomos, meio extracelular, membrana.
A composição química do RNA difere do DNA em diversos aspectos:
ele contém o açúcar ribose no lugar da desoxirribose e a base uracila (U) ao invés da timina (T); dobra-se, adquirindo
diversas formas importantes na sua função. O DNA é composto por fita dupla enquanto o RNA é composto por uma fita
simples.
As células produzem vários tipos funcionalmente de RNAs tais como o mRNA, que transporta instruções de
como fazer proteínas, tRNA que atua como molécula adaptadora de síntese de proteína e o rRNA que é um dos
componentes dos ribossomos.
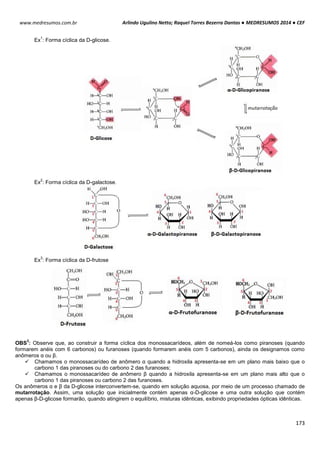
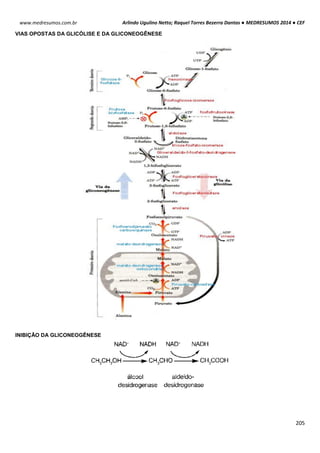
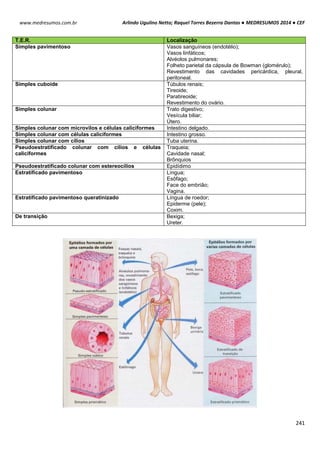
TIPOS DE RIBOSSOMOS
Os ribossomos apresentam componentes que são
designados pelos seus “valores S”, ou seja, sua taxa
sedimentação em uma ultracentrifugação. Embora os ribossomos
tanto dos eucariotos como dos procariotos apresentem
semelhança na estrutura e na funcionalidade, eles diferem no
tamanho e no número dos seus componentes proteicos.
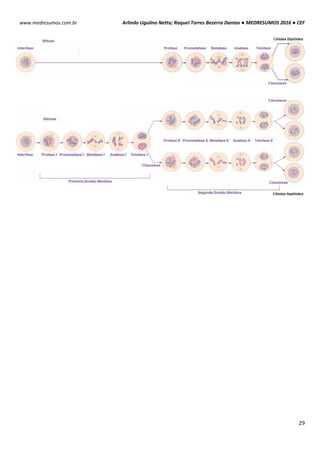
Essas estruturas são compostas por duas subunidades
uma grande e outra pequena de RNAs-ribossomais (rRNAs) que
se encaixa entre si para formar um ribossomo completo. O
ribossomos 70S procariótico é formado por uma subunidade 50S
(grande) que consiste nos rRNAs 5S e 23S de 34 proteínas e
uma subunidade 30S (pequena) constituída pelo rRNA 16S de 21
proteínas. O ribossomo 80S eucariótico contém uma subunidade
60S apresentando rRNAs 5S, 5,8S e 28S com 49 proteínas e
uma subunidade 40S tendo rRNA 18S de 33 proteínas.
OBS
1
: A subunidade maior catalisa a formação das cadeias polipeptídicas; a subunidade menor estabelece a
correspondência entre os anticódons do tRNA e os códons do mRNA.
FORMAÇÃO DOS RIBOSSOMOS
O processamento de RNAs ribossomais em células procarióticas e eucarióticas acontece de forma similar. No
procarionte Escherichia coli, por exemplo, para cada rRNA disperso no genoma existem sete operons (unidade de
expressão gênica procariótica que inclui genes estruturais coordenadamente regulados, e elementos controladores que
são reconhecido por produtos de genes reguladores) diferentes, e essa unidade contém uma cópia de cada sequência
de rRNA 5S, 16S e 23S.](https://image.slidesharecdn.com/medresumos2016-cef-170904035900/85/Medresumos-2016-cef-30-320.jpg)
























































![Arlindo Ugulino Netto; Raquel Torres Bezerra Dantas ● MEDRESUMOS 2016 ● CEF
91
www.medresumos.com.br
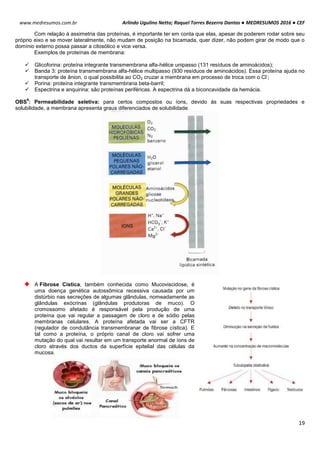
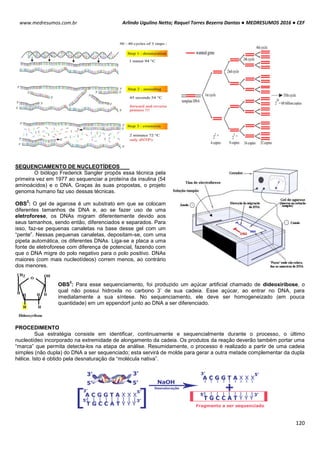
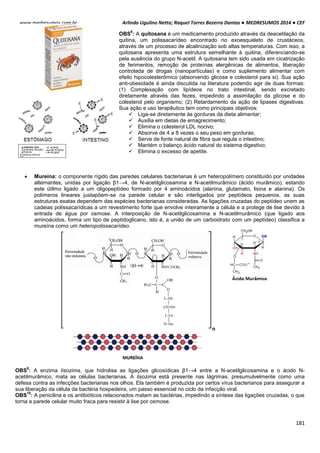
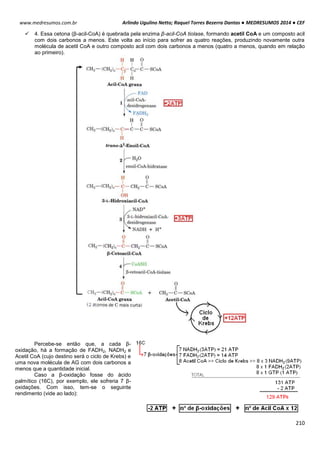
TONICIDADE DAS SOLUÇÕES
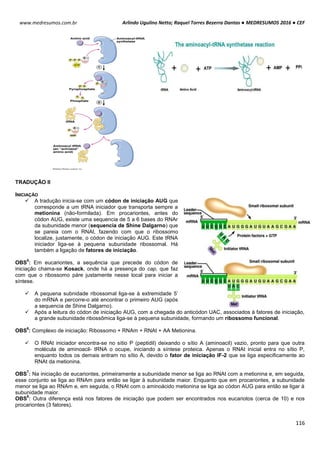
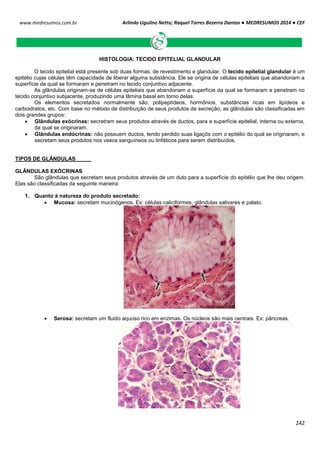
Células biológicas colocadas em diferentes soluções podem apresentar diferentes formas, dependendo da
concentração externa e da permeabilidade da membrana celular. Um exemplo clássico é feito com as hemácias, que
possuem uma concentração interna de 0,3 osm. Note como ela se comporta em diferentes situações de concentração:
Neste caso as concentrações dentro e fora da célula são iguais, mais isso não quer dizer que não há
movimentação entre a membrana celular. É importante salientar que a quantidade de H2o que entra
na célula é aproximadamente igual a sai (Iso = “igual” e Tonus = “força”).
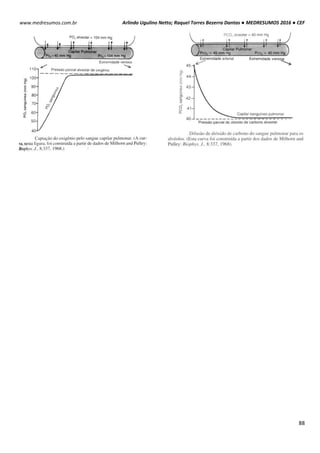
Quando uma hemácia é mergulhada em uma solução hipotônica, ou seja, a concentração de soluto
no interior da célula é maior que no meio extracelular, consequentemente o meio extracelular é mais
diluído que o intracelular. Com isso surgira uma Phid de fora para dentro promovendo a entrada de
água para o interior da hemácia. Assim a célula fica túrgida, inchada. Phid fora > Phid dentro.
HIPO = “baixo” / TONUS = “força”
Em uma solução hipertônica em que a concentração de soluto no interior da célula é baixo em
relação ao meio externo enquanto a concentração da água é maior. Com isso ocorrerá o inverso do
que acontece com o a solução hipotônica, a pressão hidrostática surgirá de dentro da célula,
havendo assim uma perda de água por parte da célula deixando-a murcha.
Intracelular: ↑ [NaCl] ↓ [H2O]
Extracelular: ↓ [NaCl] ↑ [H2O]
Solução Hipotônica célula ganha água
Intracelular: ↓ [NaCl] ↑ [H2O]
Extracelular: ↑ [NaCl] ↓ [H2O]
Hipertônica célula perde água
DISSOCIAÇÃO
1. Solutos não se dissociam: As concentrações molar e osmolar são,
evidentemente, as mesmas:
Cmolar = Cosmolar
2. Os solutos se dissociam completamente: A concentração osmolar é
igual à concentração molar multiplicada pelo número de partículas (n):
Cosm = CM x n
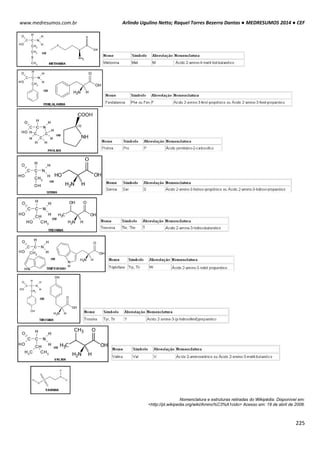
Exemplo I: Qual a concentração osmolar de NaCl 0,1 M?
NaCl libera duas partículas:
Cosm = 0,1 x 2 = 0,2 osmolar
Exemplo II: No caso do CaCl2, que libera 3 partículas, multiplicar por 3: a
concentração de partículas de CaCl2 0,15M será:
Cosm = 0,15 x 3 = 0,45](https://image.slidesharecdn.com/medresumos2016-cef-170904035900/85/Medresumos-2016-cef-91-320.jpg)






![Arlindo Ugulino Netto; Raquel Torres Bezerra Dantas ● MEDRESUMOS 2016 ● CEF
99
www.medresumos.com.br
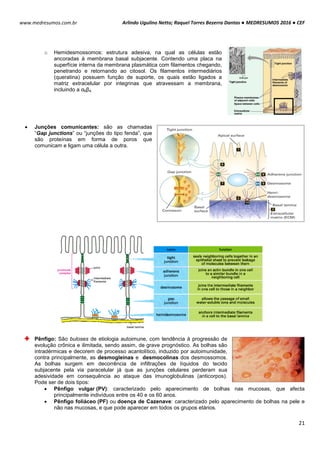
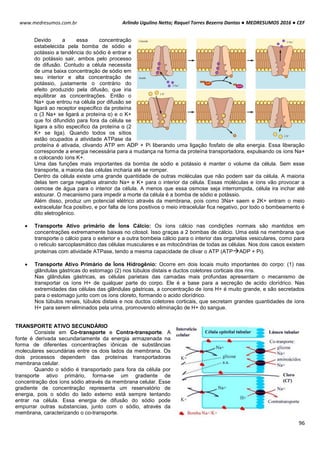
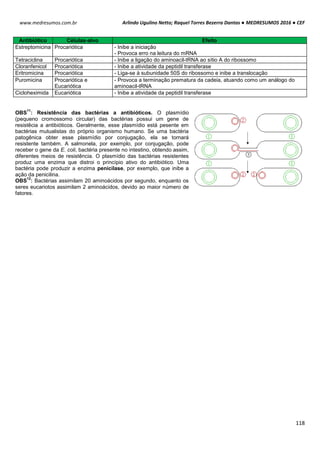
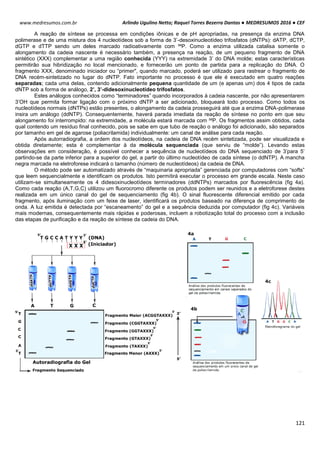
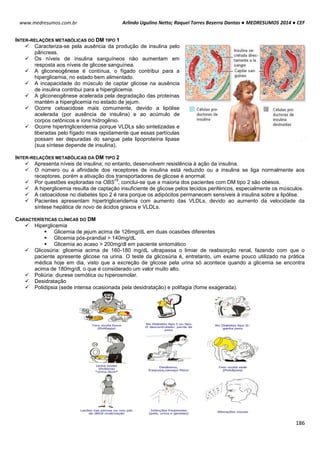
POTENCIAL DE NERST
O potencial de Nerst é determinado pela grandeza de um íon específico dentro e fora da célula (T=37ºC). Esse
potencial se opõe ao potencial de difusão – quanto maior essa proporção, maior será a tendência que esse íon se
difunda em uma direção e maior será o potencial de Nerst para impedir o potencial de difusão.
(-) se íon for positivo
(+) se o íon for negativo
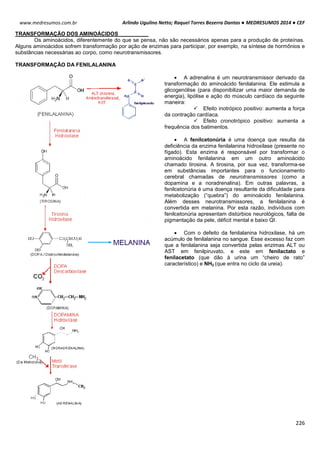
Ex: Para o K+, temos:
Vk+ = -61 . log [K+intracelular] Vk+ = -61 . log [140mEql
-1
]
[K+extracelular] [4mEql
-1
]
Vk+ = -61 . log35
Vk+ = -94mV
Ex²: Para o Na+, temos:
VNa+ = -61 . log [Na+intracelular] VNa+ = -61 . log [14mEql
-1
]
[Na+extracelular] [142mEql
-1
]
VNa+ = +61mV
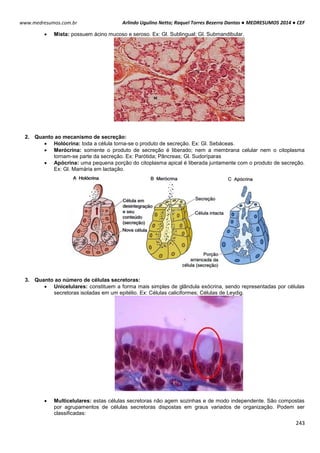
POTENCIAL DE REPOUSO / REGIME ESTACIONÁRIO / ESTADO FIXO
Esse processo tem sua origem em um mecanismo simples, de alternância entre transporte ativo e passivo de
íons.
Potencial de Difusão do K+: A membrana celular é mais permeável ao íon K+. Devido a diferença de
concentração causada pela bomba de sódio e potássio, esses íons saem da célula para o meio exterior, por
meio de canais volt-dependentes. A saída de K+ torna o meio interno da célula negativo e o externo positivo. O
potencial calculado pelo potencial de Nerst é de -94mV, ou seja, 94mV mais negativo que o meio externo.
Potencial de Difusão do Na+: A membrana de uma fibra muscular é pouco permeável ao Na+, resultando em
altas concentrações desse íon fora da célula e baixa dentro da célula, diferenca essa causada também pela
bomba de sódio e potássio. Com isso com o excesso de íon K+ que sai da célula e pouco Na+ entra, aumenta a
diferença das cargas no interior (-) e exterior (+).
Bomba de sódio e potássio: A bomba de sódio e potássio é a principal causadora da diferença de
concentração entre os íons Na+ e K+, devido ao transpore ativo de 3 íons Na+ para o exterior e 2 K+ para o
interior da célula, deixando assim um deficil de íons positivos na parte interior da célula.
Na+ (externo) 142 mEq/l K+ (externo) 4 mEq/l
Na+ (interno) 14 mEq/l K+ (interno) 140 mEq/l](https://image.slidesharecdn.com/medresumos2016-cef-170904035900/85/Medresumos-2016-cef-99-320.jpg)

















































































































![Arlindo Ugulino Netto; Raquel Torres Bezerra Dantas ● MEDRESUMOS 2014 ● CEF
231
www.medresumos.com.br
insulina. O glucagon age na conversão do ATP (trifosfato de adenosina) a AMP-cíclico, composto importante na
iniciação da glicogenólise, com imediata produção e liberação de glicose pelo fígado. Além disso, o glucagon
age nos seguintes mecanismos:
Ácidos graxos livres e cetoácidos em níveis aumentados no sangue
Produção de uréia aumentada
Estímulo da proteólise
Estímulo da lipólise: liberação de ATP (pela β-oxidação) para fornecimento de energia para realizar a
glicólise.
Albumina: é uma proteína de alto valor biológico presente na clara do ovo, no leite e no sangue. A albumina é
fundamental para a manutenção da pressão osmótica, necessária para a distribuição correta dos líquidos
corporais entre o compartimento intravascular e o extravascular, localizado entre os tecidos. Tem como funções:
manutenção da pressão osmótica; transporte de hormônios tireoideanos; transporte de hormônios lipossolúveis;
transporte de ácidos graxos livres; transporte de bilirrubina não conjugada; transporte de fármacos e drogas;
união competitiva com ions de cálcio; controle do pH. Seu excesso ocasiona diversas doenças, como problemas
renais e hepáticos. Além disso, o consumo excessivo de albumina provoca ganho de peso, sendo que um
aumento em massa muscular sem acúmulo de gorduras e também é responsável pelo fator anti-catabólico ou
seja bloqueia a perda de músculos.
Corticotropina: formado por 39 resíduos de aminoácidos. É um hormônio secretado pelo hipotálamo que
estimula o córtex da glandula adrenal à produzir hormônios (cortisol).
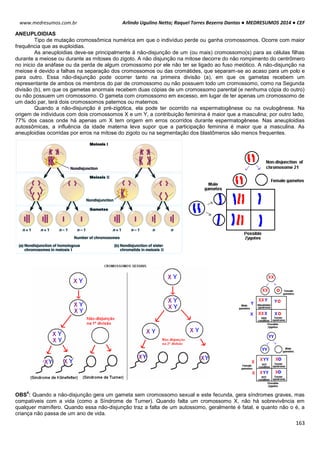
Aspartame: é um adoçante artificial utilizado para substituir o açúcar comum. É
potanto um dipeptídeo sintético formado pela fenilalanina e ácido aspartico: N-L-alfa-
aspartil-L-fenilalanina 1-metilester. Por esta razão, produtos alimentares contendo
aspartamo devem mostrar um aviso do tipo "Contém uma fonte de fenilalanina", pois
a ingestão excessiva deste aminoácido pode ser prejudicial em indíviduos com
fenilcetonúria. Ele tem maior poder de adoçar (cerca de 200 vezes mais doce que a
sacarose) e é menos denso.
Ocitocina: 9 resíduos de aminoácidos. Produzida pela hipófise posterior e estimula as contrações na hora do
parto e a liberação de leite pelas glandulas mamárias.
Encefalina: 5 resíduos de aminoácidos formados no SNC que se ligam às células do cérebro e induzem a
analgesia. A encefalina é um pentapeptídeo que termina ou com o aminoácido leucina ("Leu") ou com o
aminoácido metionina ("Met"). Ambos são produtos do gene proencefalina.
Metionia-encefalina ([Met]-encefalina) é Tyr-Gly-Gly-Phe-Met.
Leucina-encefalina ([Leu]-encefalina) é Tyr-Gly-Gly-Phe-Leu.
Escotofobina: primeira célula de memória.
Glutationa: é um poderoso antioxidante tendo como local ativo o tiol (SH) da cisteína. É um tripeptídeo formado
por GLU-CIS-ALA. Pode encontrar-se na forma reduzida (GSH) ou
oxidada (GSSG, forma dimerizada da GSH). A importância deste par
é tal que a razão GSH/GSSG é normalmente utilizada para estimar
o estado redox dos sistemas biológicos. Em situações normais a
GSSG representa apenas uma pequena fração da glutationa total
(menos de 10%). A GSH pode, no entanto, também formar
dissulfuretos do tipo GSSR com o tiol da cisteína presente em
proteína.
2GSH + H2O2 GSSG + 2H2O
2GSH + ROOH GSSG + ROH + 2H2O
GSSG + NADPH+H
+
2GSH + NADP
+
Vasopressina (ADH): hormônio antidiurético sintetizado pelo hipotálamo e armazenado na hipófise posterior
formado por 9 resíduos de aminoácidos. O alcool bloqueia a secreção de ADH por ser uma substância diurética,
fazendo com que haja uma grande excreção de água pela urina.
OBS
3
: A diabetes insipidus (DI) é uma doença caracterizada pela sede pronunciada e pela excreção de grandes
quantidades de urina muito diluída. Esta diluição não diminui quando a ingestão de líquidos é reduzida. Isto denota a
incapacidade renal de concentrar a urina. A DI é ocasionada pela deficiência do hormônio antidiurético (vasopressina) ou
pela insensibilidade dos rins a este hormônio. A diurese excessiva e a sede intensa são típicos da DI. Os sintomas da](https://image.slidesharecdn.com/medresumos2016-cef-170904035900/85/Medresumos-2016-cef-231-320.jpg)




















O documento apresenta um resumo sobre o Módulo Célula, Estrutura e Função (CEF). O módulo abrange assuntos importantes sobre a estrutura e função celular, incluindo citologia, biofísica, genética, bioquímica e histologia. Além de aulas teóricas, o módulo inclui aulas práticas em laboratório para estudar a célula. O resumo serve como guia para os estudos no módulo.



































































