Baixado 230 vezes





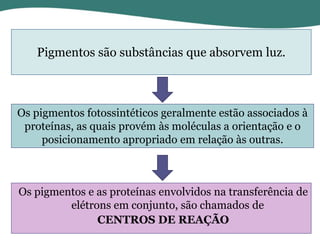
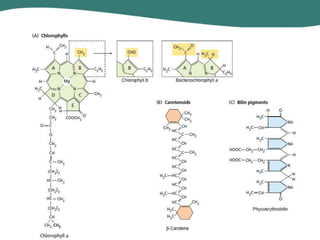
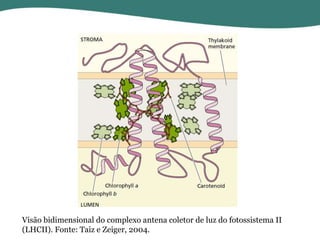
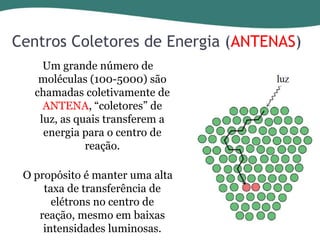

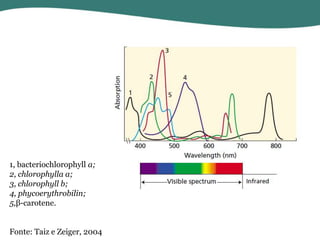
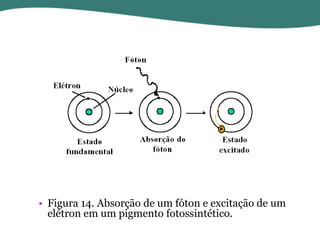
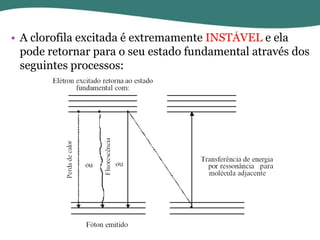
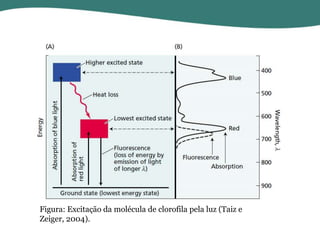
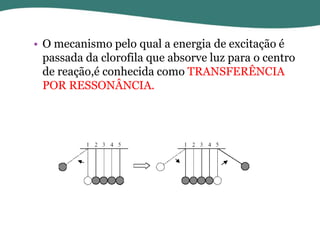
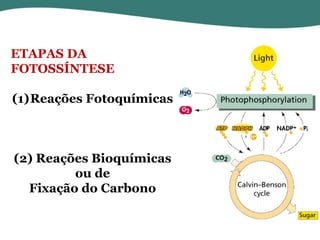
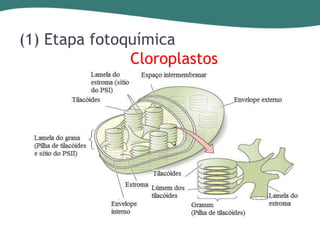
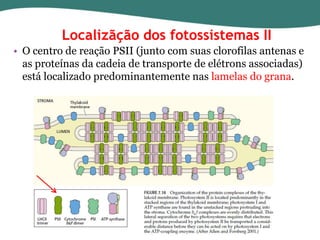
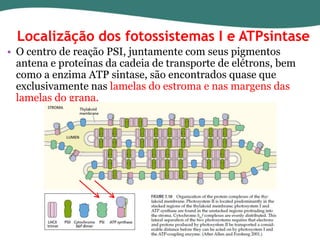
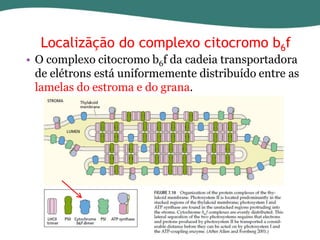
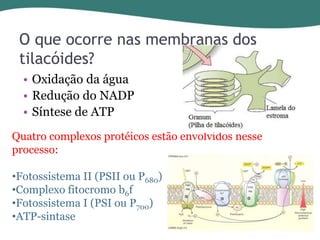
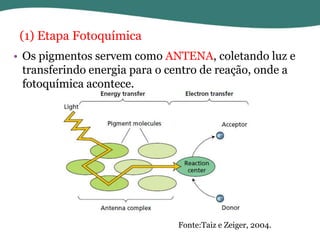
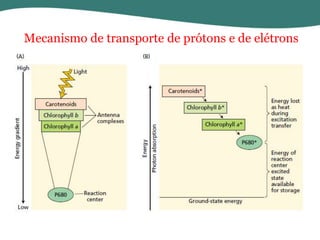
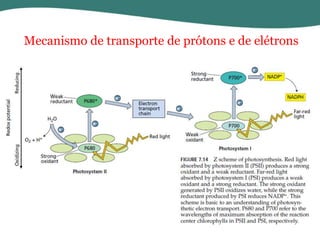
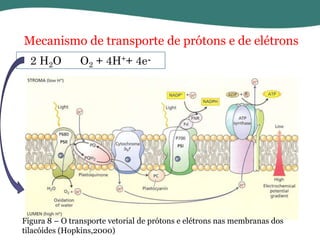
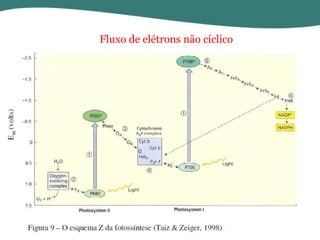
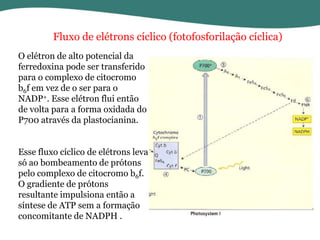


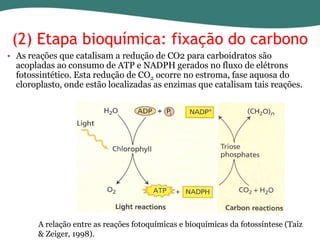
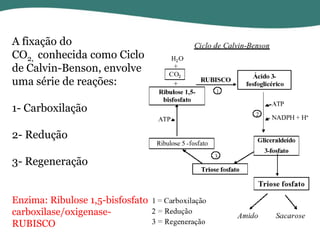
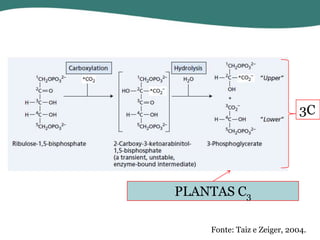
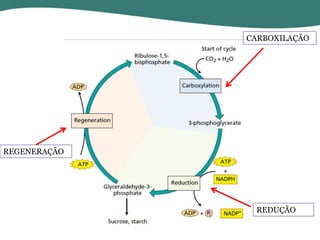
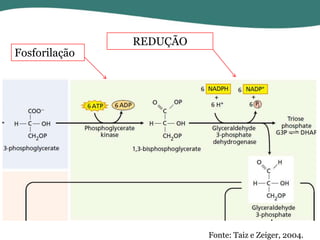
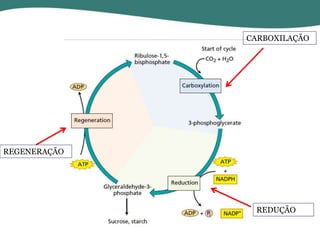


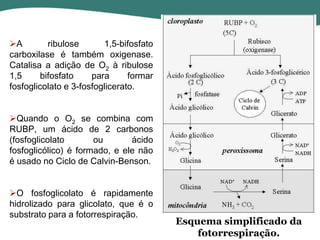

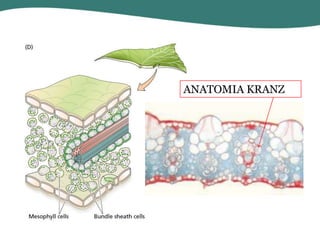
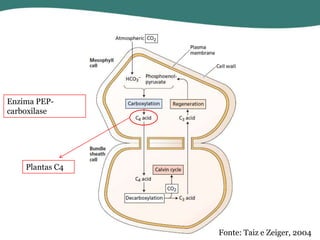
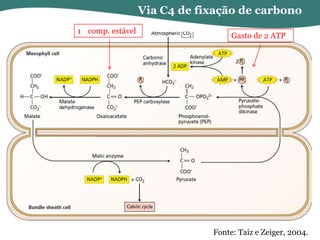



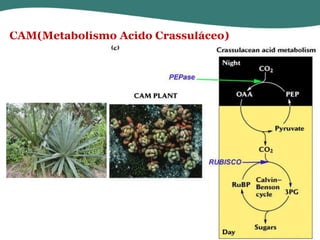
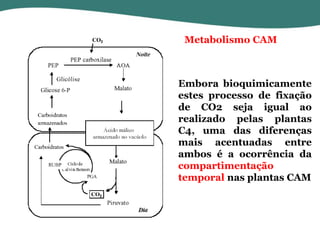
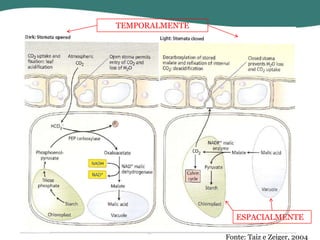
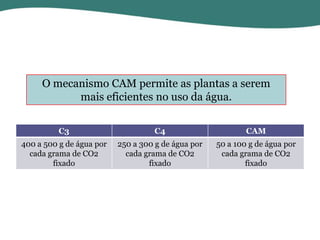



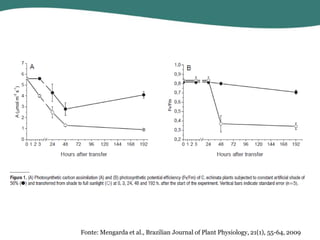



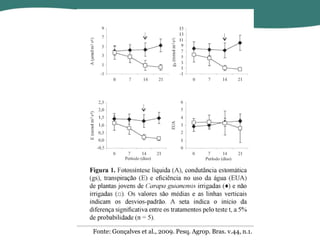

O documento discute os processos de fotossíntese em plantas. A fotossíntese converte a energia solar em energia química através da absorção da luz pelo clorofila e reações fotoquímicas e bioquímicas. As reações fotoquímicas geram NADPH, ATP e O2. As reações bioquímicas fixam o CO2 na forma de açúcares usando NADPH e ATP. Plantas C3, C4 e CAM variam nas vias de fixação do CO2.























































































