Transferir como PPSX, PPTX




























![(-) ATP
(-) NADH
(+) ADP
(-) ATP
(-) NADH
(-) SuccinilCoA
Oxalo-
acetato Citrato
Malato Isocitrato
a-ceto-
glutaratoSuccinato
Succinil-
CoA
(-) ATP
(-) NADH
(-) SuccinilCoA
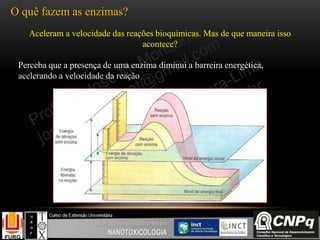
Importante:
Um alto valor da relação [ATP]/
[ADP] ou da relação
[NADH]/ [NAD+] INIBE o ciclo
de Krebs
(-) ATP
(-) NADH
(+) ADP
(-) ATP
(-) NADH
(-) SuccinilCoA
Oxalo-
acetato Citrato
Malato Isocitrato
a-ceto-
glutaratoSuccinato
Succinil-
CoA
(-) ATP
(-) NADH
(-) SuccinilCoA
Importante:
Um alto valor da relação [ATP]/
[ADP] ou da relação
[NADH]/ [NAD+] INIBE o ciclo
de Krebs
(-) ATP
(-) NADH
(-) SuccinilCoA
Oxalo-
acetato Citrato
Malato Isocitrato
a-ceto-
glutaratoSuccinato
Succinil-
CoA
(-) ATP
(-) NADH
(-) SuccinilCoA
Importante:
Um alto valor da relação [ATP]/
[ADP] ou da relação
[NADH]/ [NAD+] INIBE o ciclo
de Krebs
Oxalo-
acetato Citrato
Malato Isocitrato
a-ceto-
glutaratoSuccinato
Succinil-
CoA
(-) ATP
(-) NADH
(-) SuccinilCoA
Importante:
Um alto valor da relação [ATP]/
[ADP] ou da relação
[NADH]/ [NAD+] INIBE o ciclo
de Krebs
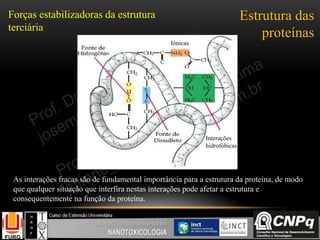
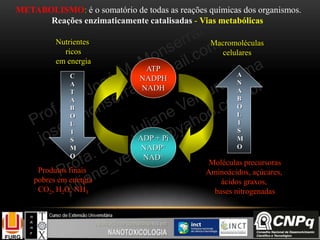
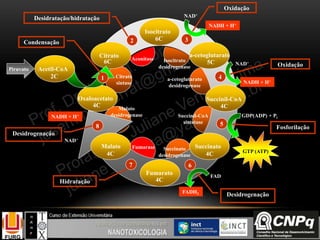
Regulação do ciclo de Krebs
A regulação acontece nas
enzimas citrato sintase, isocitrato
desidrogenase e -cetoglutarato
desidrogenase.
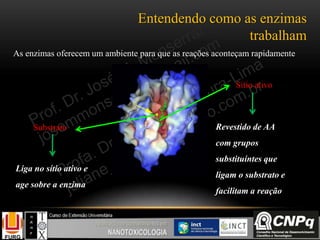
A principal função do ciclo de
Krebs é fornecer coenzimas
reduzidas com NADH e FADH2
para serem re-oxidadas na cadeia
transportadora de elétrons e
consequentemente gerar ATP pelo
processo de fosforilação oxidativa
como será visto a seguir...](https://image.slidesharecdn.com/nanobioqumica-150924182751-lva1-app6892/85/Bioquimica-aplicada-a-Nanotoxicologia-41-320.jpg)

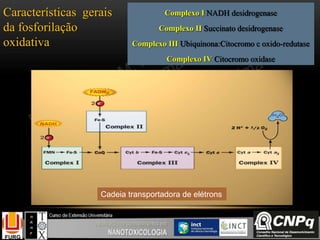

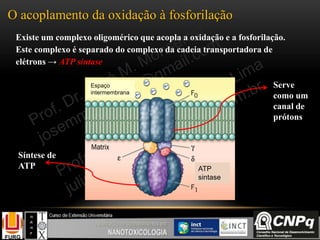

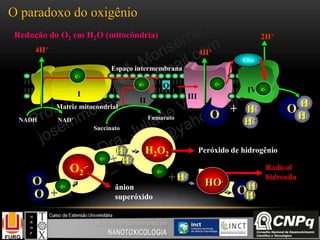


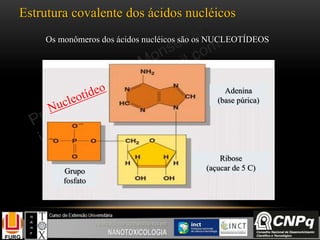
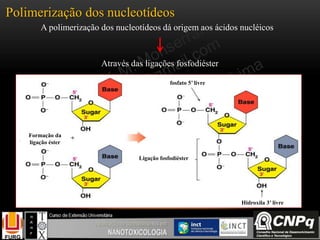




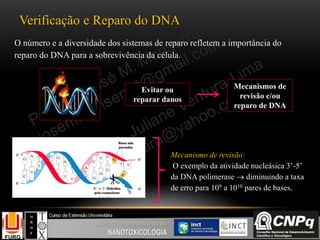
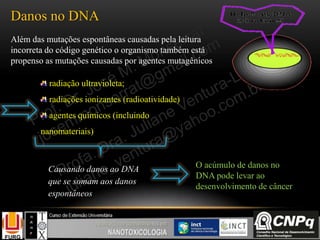
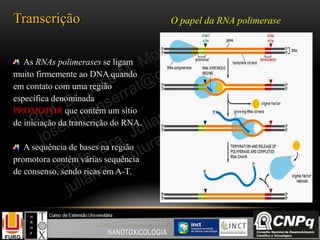
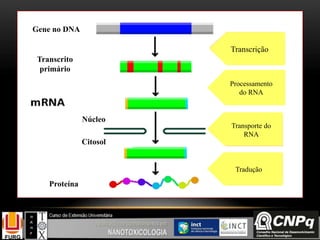

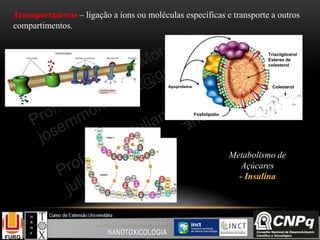
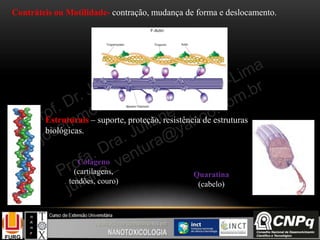
O documento descreve um módulo de bioquímica aplicada à nanotoxicologia ministrado por professores da Universidade Federal do Rio Grande. O módulo aborda tópicos como estrutura e função de proteínas, enzimas, lipídios, ciclo de Krebs, fosforilação oxidativa e estrutura de ácidos nucléicos. O documento fornece detalhes sobre a estrutura e função de proteínas e enzimas.



































































