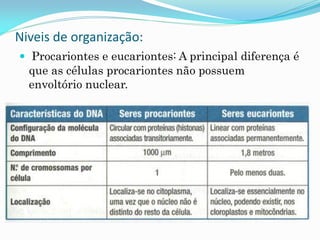
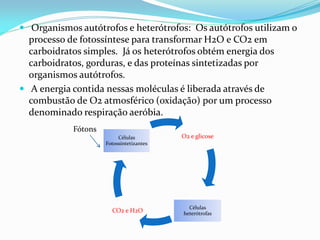
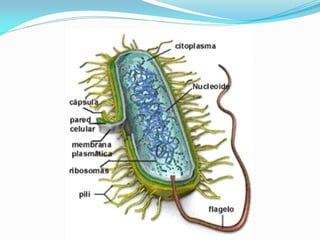
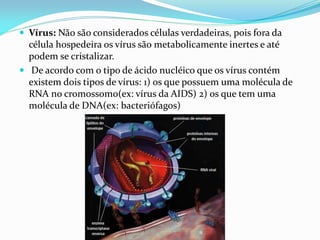
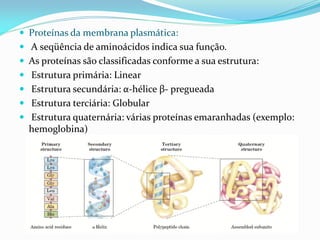
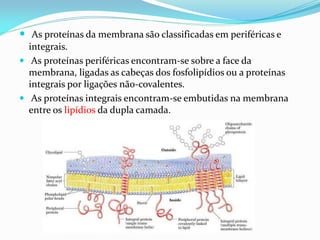
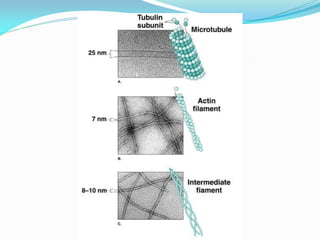
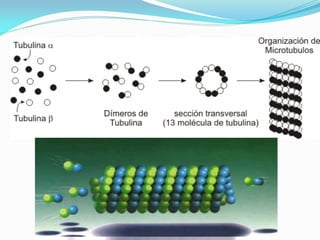
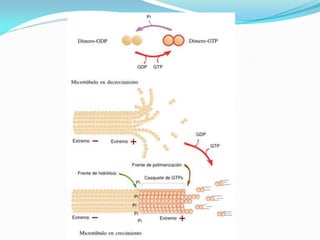
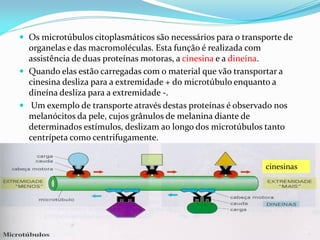
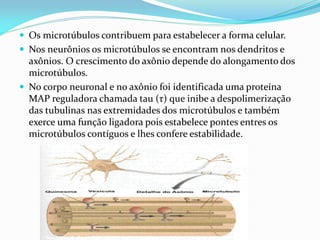
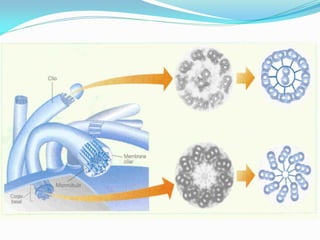
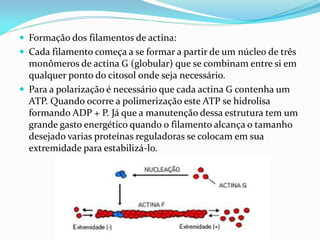
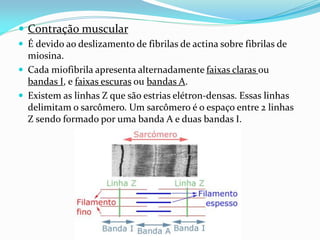
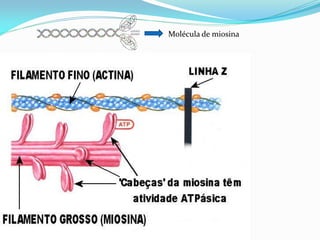
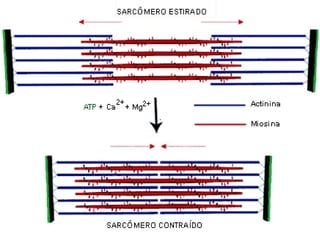
O documento discute a organização de células eucariontes e procariotos. Descreve as diferenças entre procariotos e eucariotos, organismos autótrofos e heterótrofos, e discute a estrutura e função das membranas celulares, incluindo os tipos de transporte através das membranas. Também aborda brevemente o citoesqueleto e movimentos celulares.