Baixado 497 vezes


![Copyright © Fundação de Estudos e Pesquisas Agrícolas e Florestais
1ª edição 2010
Tiragem: 500 exemplares
Capa e Projeto gráfico
Colorau Comunicação e Design
Diagramação
Diagrama Editorial
FEPAF
Fundação de Estudos e Pesquisas Agrícolas e Florestais
Unesp Campus de Botucatu Lageado.
Fazenda Experimental Lageado s/nº
18.603-970 Botucatu SP Brasil
Tel.: (14) 3882-7373 fepaf@fca.unesp.br
www.fepaf.org.br
FICHA CATALOGRÁFICA ELABORADA PELA SEÇÃO TÉCNICA DE AQUISIÇÃO E TRATAMENTO
DA INFORMAÇÃO – SERVIÇO TÉCNICO DE BIBLIOTECA E DOCUMENTAÇÃO - UNESP - FCA
- LAGEADO - BOTUCATU (SP)
T764
Tópicos em ecofisiologia da cana-de-açúcar / Editores
técnicos: Carlos Alexandre Costa Crusciol ... [et
al.]. – Botucatu: FEPAF - Fundação de Estudos e Pesquisas Agrícolas e Florestais, 2010
111 p. : il., gráfs., tabs., fots. color.
ISBN 978-85-98187-25-9
1. Cana-de-açúcar. 2. Ecofisiologia. I. Crusciol, Carlos Alexandre Costa. II. Silva, Marcelo de Almeida. III.
Rossetto, Raffaella. IV. Soratto, Rogério Peres. V. Fundação de Estudos e Pesquisas Agrícolas e Florestais. VI.
Título.
CDD 21.ed. (633.61)
iniciais.indd 2
14/10/10 09:38](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-2-320.jpg)






























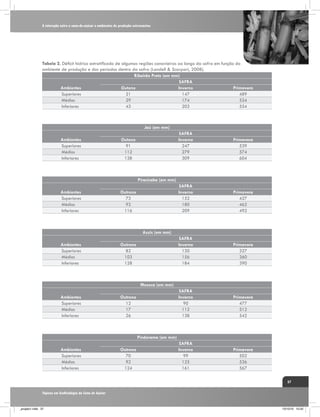

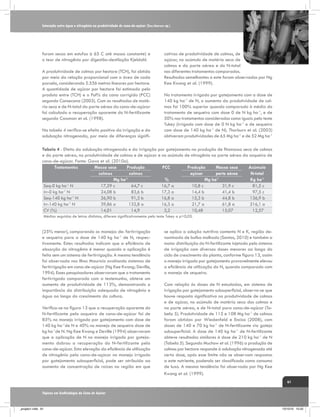
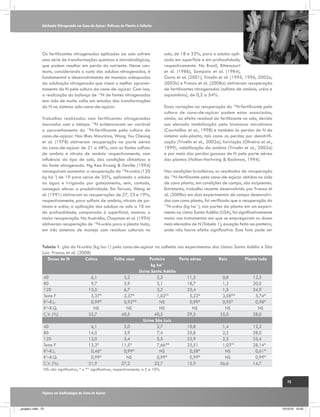
![A interação entre a cana-de-açúcar e ambientes de produção estressantes
Tabela 3. Quantificação biométrica utilizados pelo IAC em ensaios de competição varietal em regiões
de déficits hídricos contrastantes.
Altura Colmo (cm)
Diâmetro colmo (cm)
Perfilhamento (número/m)
Variedade
FAV.
DESF.
ISh
FAV.
DESF.
ISh
FAV.
DESF.
ISh
IAC911155
240
233
0,97
2,3
2,2
0,96
10,5
13,4
1,28
IACSP931042
270
261
0,97
2,9
2,4
0,83
10,2
10,3
1,01
IACSP941104
269
251
0,93
2,9
2,3
0,79
12,3
9,9
0,80
IACSP951218
246
270
1,10
3,0
2,5
0,83
11,0
10,3
0,74
IACSP952078
250
227
0,91
2,6
2,1
0,81
12,6
12,8
1,02
IACSP955094
265
246
0,93
2,6
2,5
0,96
11,5
11,4
0,99
IACSP962037
265
264
1,00
2,9
2,6
0,90
10,3
10,2
0,99
IACSP962041
252
284
1,13
2,7
2,4
0,89
9,9
10,6
1,07
IACSP962042
285
280
0,98
2,7
2,6
0,96
11,0
11,3
1,03
IACSP963057
275
257
0,93
2,9
2,2
0,76
9,5
10,3
1,08
IACSP963060
261
255
0,98
3,0
2,5
0,83
10,8
10,8
1,00
IACSP963069
253
261
1,03
2,9
2,5
0,86
10,7
10,4
0,97
IACSP976645
249
256
1,03
2,6
2,5
0,96
11,0
9,9
0,90
IACSP976671
259
279
1,08
2,7
2,4
0,89
9,7
11,0
1,13
IACSP976682
256
263
1,03
2,9
2,4
0,83
10,7
10,8
1,01
RB72454
247
290
1,17
2,9
2,3
0,79
9,7
8,3
0,86
RB867515
274
315
1,15
3,0
2,5
0,83
11,6
11,8
1,02
SP832847
236
304
1,29
2,5
2,2
0,88
11,3
9,5
0,84
SP9642
278
250
0,90
2,8
2,3
0,82
11,1
11,9
1,07
MÉDIAS
260
267
1,03
2,7
2,4
0,86
10,9
10,6
1,00
MÉDIAS PAD.
261
288
2,7
2,3
11,3
10,1
DMS (10%)
78
61
0,5
0,4
3,1
3,5
CV
10,38
7,92
6,77
5,35
10,02
11,48
4.2. Parâmetros fisiológicos
Trocas gasosas e atividade fotoquímica
As trocas gasosas são avaliadas com um analisador de
gases por infravermelho modelo Li-6400F (Licor, EUA),
com fluorômetro acoplado. As variáveis de trocas gasosas estudadas são: assimilação de CO2 (PN, μmol m-2 s-1);
transpiração (E, mmol m-2 s-1); condutância estomática (gS,
mol m-2 s-1); e concentração intercelular de CO2 (CI, μmol
mol-1). A diferença de pressão de vapor entre a folha e
o ar (DPVL, kPa) e a temperatura foliar (TF, ºC) também
são monitoradas com o Li-6400. Com o Li-6400 é possível
o cálculo da eficiência instantânea de carboxilação (PN/
CI), assim como a eficiência intrínseca do uso da água
(EUA = PN/gS).
I
As variáveis da atividade fotoquímica avaliadas são: a
fluorescência mínima (FO) e máxima (FM) foram monitoradas em tecidos foliares adaptados ao escuro (30 min).
A fluorescência instantânea (F’S) e máxima (F’M) foram
avaliadas em tecidos adaptados à luz. Os valores de FM
e F’M foram determinados após um pulso de saturação e
a fluorescência mínima após a excitação do fotossistema
I (F’O) com luz vermelha distante. A partir dos valores de
FO, FM, F’S e F’M, foram calculadas a fluorescência variável no escuro (FV=FM-FO) e no claro (_F=F’M-F’S), sendo
’
possível a obtenção de algumas variáveis fotoquímicas:
eficiência quântica potencial (FV/FM) e efetiva (_F’/F’M)
do fotossistema II (FSII); coeficiente de extinção não
fotoquímica da fluorescência [NPQ=(FM-F’M)/F’M]; e o
transporte aparente de elétrons [ETR’S = Q x _F/F’M x
0,4 x 0,85] (McCormick et al., 2008).
Potencial da água na folha
O estado hídrico das plantas é avaliado pelo potencial
da água na folha, medido na antemanhã com o auxílio
de uma câmara de pressão modelo 3005 (Soilmoisture
Equip., EUA).
39
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 39
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-39-320.jpg)







![Alterações morfofisiológicas na cana-de-açúcar em resposta à deficiência hídrica
stress effect on sugarcane photosynthesis. Proccedings
South African Sugar Technology Association, v. 80,
p. 148-158, 2006.
LARCHER, W. Ecofisiologia Vegetal. São Carlos: RIMA,
2000. 531p.
MACHADO, R.S. Respostas fisiológica de genótipos
de cana-de-açúcar ao déficit hídrico imposto na fase
inicial de desenvolvimento. Campinas: IAC, 2009. 64p.
Dissertação (Mestrado).
PAIVA, R.; OLIVEIRA, L.M. Fisiologia e Produção Vegetal.
Lavras: Editora UFLA, 2006. 104p.
PEREIRA, P.G.; BERGAMASCHI, H.; ROSA, L.M.G.; FRANÇA, S. Efeitos de diferentes níveis de déficit hídrico na
fotossíntese e condutância foliar em milho. Revista Brasileira de Agrometeorologia, v.11, n.1, p. 53-62, 2003.
PIMENTEL, C. Metabolismo de carbono na agricultura
tropical. Seropédica: EDUR, 1998. 150p.
SCARPARI, M.S. PREDPOL: um modelo de previsão da
maturação da cana-de-açúcar visando planejamento
otimizado. Piracicaba:ESALQ/USP, 2007.120p. Tese
(Doutorado).
SCARPARI, M.S.; BEAUCLAIR, E.G.F. The sugarcane maturity estimation through edaphic-climatic parameters.
Scientia Agricola, v.61, n.5, p.486-491, 2004.
SEGATO, S.V.; MATTIUZ, C.F.M.; MAZAMBANI, A.E. Aspectos fenológicos da cana-de-açúcar. In: Atualização
em produção de cana-de-açúcar. SEGATO, S. V. [et al.].
Piracicaba: CP 2, 2006. 415p.
SHIGAKI, F.; FREITAS, N.; BERTO, A.; CEDDIA, M.B.;
ZONTA, E.; LIMA, E. Influência do estresse hídrico nos
parâmetros de crescimento, acúmulo e produtividade de
diferentes variedades de cana-de-açúcar em Miracema –
RJ. Revista Universidade Rural: Série Ciências da Vida,
v.24, n.1, p.63-71, 2004.
PIMENTEL, C. A relação da planta com a água. Seropédica: EDUR, 2004. 191p.
SILVA, M.A.; JIFON, J.L.; DA SILVA, J.A.G.; SHARMA, V.
Use of physiological parameters as fast tools to screen
for drought tolerance in sugarcane. Brazilian Journal of
Plant Physiology, v. 19, n. 3, p. 193-201, 2007.
RAO, C.M.; KUMAR, M.V.; REDDY, L.K. Management
of sugarcane clones under limited moisture situations
(drought) at formative stage. Indian Sugar, february, p.
949-953, 2005.
SILVA, M.A.; DA SILVA, J.A.G.; ENCISO, J.; SHARMA,
V.; JIFON, J. Yield components as indicators of drought
tolerance of sugarcane. Scientia Agricola, v. 65, n. 6, p.
620-627, 2008.
ROBERTSON, M.J.; INMAN-BAMBER, N.G.; MUCHOW,
R.C. WOOD, A.W. Physiology and productivity of sugarcane with early and mid-season water deficit. Field
Crops Research, v. 64, p. 211-227, 1999.
SILVA, C.R. Evapotranspiração e desenvolvimento de limeira ácida ‘Tahiti’ na ausência e presença de estresse
hídrico. Piracicaba: ESALQ/USP, 2005. Tese (Doutorado).
SAUSEN, T.L. Respostas fisiológicas de Ricinus communis
à redução na disponibilidade de água no solo.Porto
Alegre: Universidade Federal do Rio Grande do Sul/
Faculdade de Agronomia Porto Alegre, 2007.61p. Dissertação (Mestrado).
SANTOS, M.A.L. Irrigação suplementar da cana-deaçúcar (Saccharum spp.): um modelo de análise de
decisão para o Estado de Alagoas. Piracicaba: ESALQ/
USP, 2005. 101p. Tese – Doutorado.
SANTOS, R.F.; CARLESSO, R. Déficit hídrico e os processos
morfológicos e fisiológicos das plantas. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 2, n. 3, p.
287-294, 1998.
SASSAKI, R.M.; MACHADO, E.C. Trocas gasosas e condutância estomática em duas espécies de trigo em diferentes teores de água no solo. Pesquisa Agropecuária
Brasileira, v.34, n.9, p.1571-1579, 1999.
SMIT, M.A.; SINGELS, A. The response of sugarcane
canopy development to water stress. Field Crops Rese�����������������
arch, v. 98, p.91-97, 2006.
STEUDLE, E. Water uptake by roots: effects of water
déficit. Journal of Experimental Botany, v. 51, n. 350,
p. 1531-1542, 2000.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3ed. Porto Alegre:
Artmed, 2004. 719p.
VENKATARAMANA, S.; RAO, P.N.G.; NAIDO, K.M. The
effects of water stress during the formative phase on
stomatal resistence and leaf water potential and relationship with yield in tem sugarcane varieties. Field Crops
Research, v. 13, p.345-353, 1986.
VERSLUES, P.E.; AGARWAL, M.; KATIYAR-AGARWAL, S.;
ZHU, J.; ZHU, J.K. Methods and concepts in quantifying
resistance to drought,salt and freezing, abiotic stresses
that affect plant water status. The Plant Journal, v. 45,
p. 523–539, 2006.
48
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 48
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-48-320.jpg)

![Interação entre água e nitrogênio na produtividade de cana-de-açúcar (Saccharum sp.)
Estudos realizados por vários pesquisadores demonstraram que genótipos de cana-de-açúcar respondem
diferentemente ao aumento da disponibilidade hídrica
(Ramesh; Mahadevaswamy, 2000; Inman-Bamber; Smith,
2005; Smit; Singels, 2006; Gava et al., 2008). Segundo
Inman-Bamber e Smith (2005), a eficiência de utilização
de água da cana-de-açúcar está relacionada com fatores fisiológicos como: capacidade de discriminação do
isótopo estável 13C pela enzima ribulose-1,5-bifosfato; a
condutância estomática; a partição de fotoassimilados;
entre outros, e morfológicos como: índice de área foliar;
a elongação dos colmos; a partição de matéria seca;
entre outros.
A deficiência de água (estresse hídrico) provoca a
redução de trocas gasosas especialmente de CO2 e
sua condução para as folhas, reduzindo a produção
de fotassimilados (Farquhar et al., 1988; Lopes, 1988;
Inman-Bamber; Smith, 2005); diminui a área foliar e
conseqüentemente o índice de área foliar (Inman-Bamber
e Smith, 2005; Smit e Singels, 2006; Farias et al., 2008),
índice que é associado à quantidade de luz absorvida
e à fotossíntese total da cana-de-açúcar (Irvine, 1975);
provoca a redução no surgimento de novas folhas e o
aumento da abscisão foliar (Smit; Singels, 2006), devido
a elevação da concentração de ácido abscísico na planta
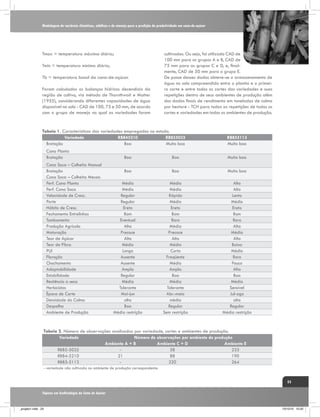
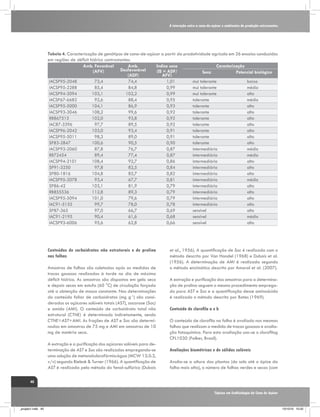
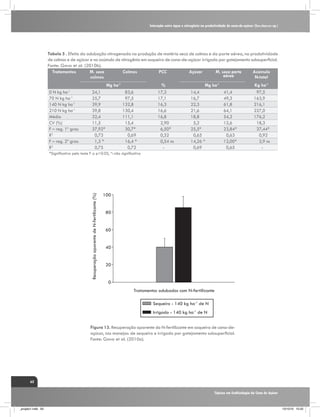
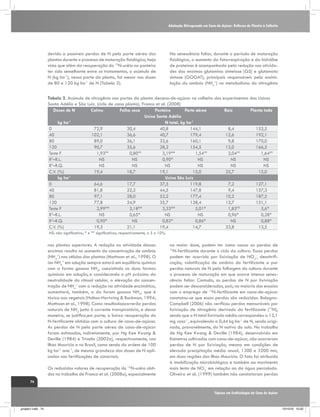
(Kramer, 1983), conforme verificado na figura 1. Assim, o
CO2 Taxa de assimilação
E
E
S
R
Fotorespiração
E
S
Custo de elétrons
para fotossíntese
H
CO2
Intercelular
R
I
R
E
S
S
E
H
Í
D
T
Clorose e indução
e senescência
S
E
S
Mudanças na
anatomia e na
estrutura foliar
T
Í
Atividade da
nitrato redutase
D
R
"Vazamento"
I
[ATP]
C
O
C
O
Condutância
estomática
Atividade
enzimas ciclo C4
Atividade
enzimas ciclo C3
ESTRESSE HÍDRICO
Fonte: Ghannoum, O. (2009),
Adaptado por: Gava, G. J. C. (2009).
Figura 1. Resumo dos principais efeitos da deficiência hídrica sobre os parâmetros fotossintéticos das folhas
de plantas de ciclo C4. Fatores correlacionados com os estômatos são indicados por linhas tracejadas e fatores
não correlacionados com linhas contínuas. O sinal (-) indica um efeito negativo. O termo “vazamento” (Φ) é
definido como a fração de CO2 fixado pela PEPC que se perde dos feixes vasculares. Fonte: Ghannoum, (2009).
50
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 50
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-50-320.jpg)
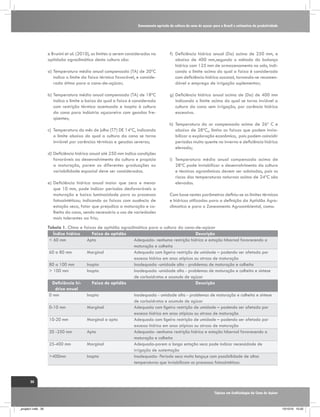
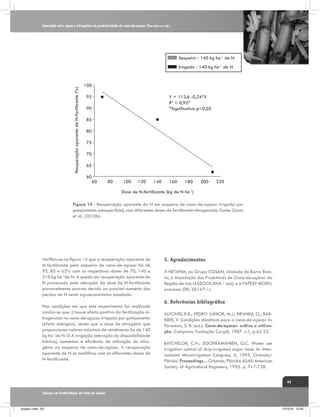
![Interação entre água e nitrogênio na produtividade de cana-de-açúcar (Saccharum sp.)
se com plantas de ciclo C3 (Figura 2).
Envolve a difusão de CO2 através de
estômatos, a sua dissolução e conversão
para HCO3-, com posterior fixação em
oxaloacetato pela enzima fosfoenol piruvato-carboxilase
(PEPC). Em última instância, o CO2 liberado no feixe da
bainha vascular das células é re-fixado pela Rubisco
(Figura 2). Como conseqüência, o fracionamento dos
isótopos de carbono possuem um valor menor em plantas
de ciclo C4 (cerca de 4 ‰) do que em plantas de ciclo
C3 (cerca de 20 ‰), (Farquhar et al, 1989). A Variação
da discriminação isotópica de carbono em plantas C4
não depende somente pi/pa (como em plantas C3), mas
também do “leakiness” (vazamento) Φ, a proporção
de CO2 liberado no feixe da bainha que não é fixado
pela Rubisco e “vaza” ou retorna de volta para o mesófilo (Farquhar, 1983; Farquhar et al., 1989; Henderson
et al., 1992).
manejo racional da água na cultura de cana-de-açúcar,
é de grande importância para a maximização da produção (Dantas Neto et al., 2006).
A deficiência de água em culturas sob condições de
campo é difícil de se avaliar porque u ma determinada
quantidade de água que entra no agroecossistema nem
sempre é aproveitada pela planta, podendo ser perdida
por vários mecanismos, como evaporação, escorrimento
superficial e lixiviação. Assim, um possível meio de se
avaliar o estresse hídrico em plantas é a determinação
do fracionamento isotópico entre os isótopos estáveis de
carbono (12C e 13C) nas folhas de plantas (Smeltekop
et al., 2002).
O fracionamento isotópico ocorre porque existem na
natureza duas formas estáveis do carbono, uma destas
formas é mais rara o 13C (ocorrendo a uma taxa de
1,1% do carbono total existente) em relação à outra, o
12
C, que são fixadas nos tecidos das plantas de forma
diferenciada (Farquhar et al, 1989).
O modelo conceitual do fracionamento isotópico do
carbono em plantas de ciclo C4 é representado pela
equação (01), desenvolvida por Henderson et al. (1992):
Os processos biológicos e físicos “preferem” compostos
contendo isótopos mais leves, por serem mais reativos
(leves), e “discriminam” os mais pesados. O 12CO2 difunde
mais rapidamente do que 13CO2 na câmara estomática
e assim enzimas reagem mais facilmente com 12CO2, em
condições de ambiente não estressante (Figura 2).
Δ = a + [b4 + Φ (b3 - s) - a] pi/pa
Nesta equação o valor (a) significa fracionamento na
difusão do CO2 atmosférico pelo estômato, como nas
plantas C3, portanto o valor a é o mesmo, em torno de
4,4‰; (b3) é o fracionamento que ocorre na carboxilação da RUBISCO (29‰); b4 significa o fracionamento
durante a dissolução e conversão para HCO3-, e fixação
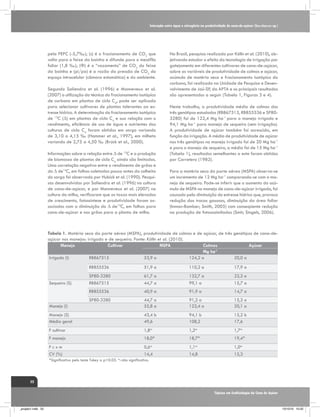
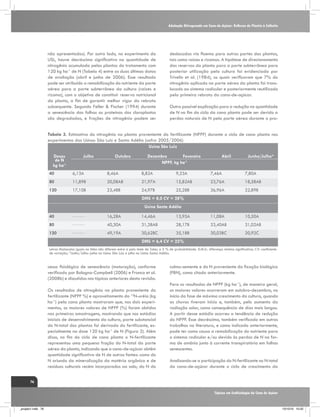
O fracionamento isotópico de carbono em plantas de
cana-de-açúcar (ciclo C4) é mais complexo comparando-
Plantas de ciclo C4
Plantas de ciclo C3
Células do mesófilo
Células do mesófilo
áHCO3-mal~1.002
PEP
Carboxilase
áCO2-CH2O=1.029
CO2
Rubisco
12
Ribulose
Bifosfato
CO2
13
Ciclo de
Calvin
[1]
H12CO3H13CO3-
Malato
Feixe de bainha vascular
áCO2-CH2O=1.029
CO2
12
Rubisco
Ribulose
Bifosfato
CO2
13
Ciclo de
Calvin
PEP
Piruvato
Triose Fosfato
Vazamento
Sacarose
Triose Fosfato
Sacarose
Produzido por:
Gava, G. J. C. (2009)
Figura 2. Representação esquemática do fracionamento isotópico de carbono na fotossíntese em plantas de ciclo C3 e C4.
51
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 51
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-51-320.jpg)
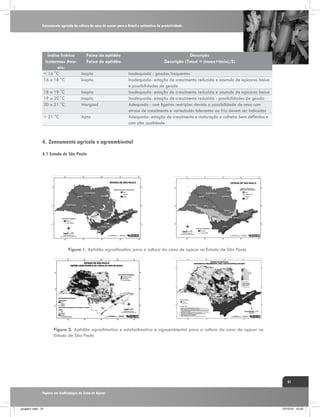
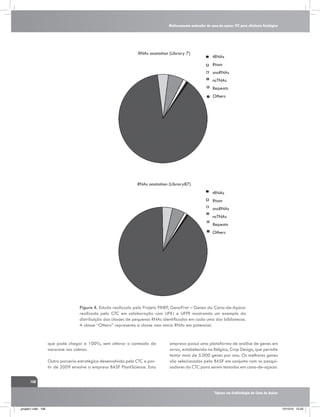
![Interação entre água e nitrogênio na produtividade de cana-de-açúcar (Saccharum sp.)
5,40
5,25
Delta 13C(% )
5,10
a
a
4,95
4,80
4,65
4,50
4,35
a
Irrigado
Sequeiro
a a
a
a
a a
b
b
4,20
4,05
3,90
C1
C2
240 DAC
C3
C1
Cultivares
C2
370 DAC
C3
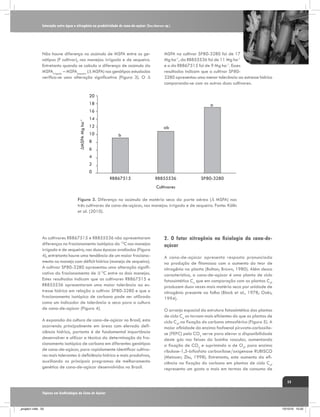
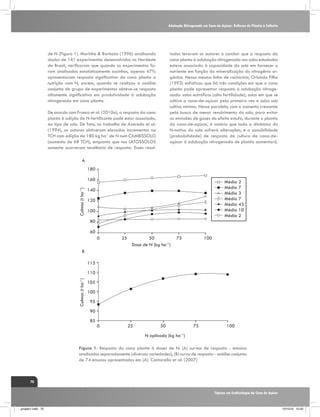
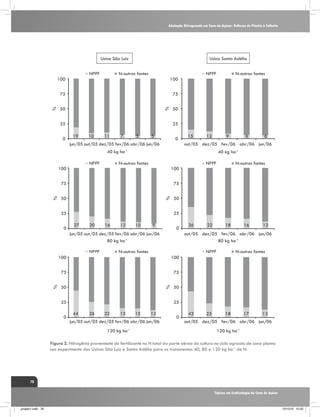
Figura 4. Fracionamento isotópico de carbono em folhas+1 de
três genótipos de cana-de-açúcar em duas épocas de amostragem
240 dias após o corte (DAC) e 370 DAC, nos manejos: irrigado
por gotejamento e de sequeiro. C1: cultivar RB867515, C2:
RB855536,e C3: SP80-3280. Fonte: Kölln et al. (2010).
Feixes da Bainha
Vascular
[CO2]M ≈ 220 µbar
CO2
C3
[CO2]FBV > 1000 µbar
CO2
C3
C4
Mesófilo
CO2
HCO3
[CO2]atm ≈ 380 µbar
Figura 5 . Representação simplificada da fixação de carbono atmosférico (fotossíntese)
de plantas de ciclo C3 e C4. Observa-se que as plantas de ciclo C4 possuem uma pressão
intercelular de CO2 maior (1.000 µbar de CO2) do que as plantas de ciclo C3 (220 µbar
de CO2). Fonte: Ghannoum, (2009).
54
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 54
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-54-320.jpg)


























![Herbicides as Stimulators, Regulators, and Ripeners
Herbicides as Stimulators, Regulators, and Ripeners
Ryan P. Viator, Caleb. D. Dalley and Edward P. Richard, Jr.
USDA-ARS Sugarcane Research Unit
The use of plant growth regulators to increase sucrose
levels in sugarcane prior to harvest (ripen) is an important
for the profitable and sustainable production of sugarcane
in the U.S. as well as in many other sugarcane industries
around the world. In most industries, harvest begins before
sucrose reaches economically profitable levels for the
grower and miller. This is especially true in the temperate
climate of the Louisiana sugarcane industry where the
window for harvesting is limited due to the risk of freezing
temperatures during the latter part of the harvest season
in December (Dalley and Richard, 2010). This paper will
discuss several ripener studies conducted by scientists at
the USDA-ARS Sugarcane Research Unit in Houma, LA.
(top, middle, and bottom). Increases of 5 to 16% in TRS
levels occurred in all sections of the stalk, but the greatest
increase (29 to 94%) occurred in the top two-thirds of
the stalk where sucrose accumulation (ripening) was still
occurring at the time of the glyphosate application.
Increases in TRS were obtained with all application timings
and sampling intervals except the November application
date. At 7 WAT, the August, September, and October
applications increased sucrose by 21%, 14%, and 8%,
respectively, compared to a no-ripener control with the
magnitude of increases in the top two-thirds of the stalk
decreasing as the application date was delayed and
natural ripening was occurring.
In the U.S., the only sugarcane ripener registered for use
is glyphosate [potassium salt of N-(phosphonomethyl)
glycine]. Glyphosate, the most widely used herbicide
worldwide (Duke and Powles, 2008), is applied at nonherbicidal rates (0.14-0.28 kg ae/ha) four to seven
weeks prior to harvest. Under favorable conditions, these
ripener applications can increase sucrose concentrations
in harvested cane by 10 to 20% (Dalley and Richard,
2010). Initially, labeled applications began in late-August
and were limited to the last and lowest yielding ratoon
crop (second-ratoon) which is harvested at the start of
the harvest season typically late-September or early
October five to six weeks before natural ripening reaches
its peak in Louisiana (USDA-ARS, 2009). Currently,
labeled applications have been modified to include all
ratoon crops in a production cycle and harvests through
November and early December.
Viator et al. (2010) conducted a similar study investigating
late-season ripener applications. The specific objectives
of this experiment were to determine if: 1) late-season
(Nov. 1) glyphosate ripener application increases sucrose
yield in four common commercial sugarcane varieties
grown in Louisiana, 2) varietal differences in susceptibility
to injury occurs in the subsequent ratoon crop from
carryover glyphosate applied to the previous ratoon, and
3) the stress on the subsequent ratoon crop associated
with glyphosate carryover from the application to the
previous ratoon crop is compounded by the retention
of post-harvest straw generated during the green-cane
harvesting of previous crop . Glyphosate was applied
on Nov. 1 (0.21 kg ae/ha) to the first-ratoon crop of the
following varieties: ‘LCP 85-384’, ‘Ho 95-988’, ‘L 97128’, and ‘HoCP 96-540’. Two weeks after the first ratoon
was harvested, the blanket of straw was either allowed
to remain or completely removed by burning. Regardless
of variety, late-season ripener application of glyphosate
to first-ratoon crops offered no increase in sucrose yield
(kg/ha). In addition, the residual herbicidal effect of
glyphosate on the sucrose yield of the subsequent secondratoon crop was not variety specific. Carryover effect of
glyphosate, when post-harvest straw was not removed,
reduced sucrose yields (kg/ha) by 13% relative to the
non-treated control as an average of the four varieties
evaluated. Sucrose yield in the second-ratoon crop was
not reduced when glyphosate was applied the prior year
and the straw was removed. The carryover effect of the
prior year’s application of glyphosate where the straw
was not removed was primarily seen as a reduction in stalk
population and photosynthesis. Current recommendations
suggest that Louisiana producers not apply glyphosate as
a ripener where the post-harvest blanket of straw cannot
be removed prior to the start of the next production year
of a crop cycle. Moreover, late-season glyphosate ripener
The application of a low dose of glyphosate as a ripener
to sugarcane that will be harvested in the last third
of Louisiana’s 100-day harvest season, which would
typically be in December, is sometimes conducted even
while its use may have little or no economic value and has
potential to cause injury to the subsequent ratoon crop
(Legendre, 2009). Richard and Dalley (2009) conducted
a study to determine the effects of different application
dates (early, mid, and late-season) on ripener efficacy.
Glyphosate, formulated with surfactant, was applied at
0.21 kg ae/ha in the first week of August, September,
October, and November of 2006 and 2007 to first-ratoon
crops of the variety ‘HoCP 96-540’. The variety HoCP 96540 is currently grown on 50% of the acreage in Louisiana
(Legendre and Gravois, 2009). Sugarcane was handharvested at 4, 5, 6, and 7 weeks after treatment (WAT)
and theoretical recoverable sucrose (TRS) was determined
for the total stalks and by dividing the stalks into thirds
89
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 89
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-89-320.jpg)
![Herbicides as Stimulators, Regulators, and Ripeners
applications are also not recommended due to a general
lack of response by sugarcane coupled with the potential
of carryover herbicidal effects caused by translocation
of glyphosate into the crown. The latter is especially
recommended against if the blanket of straw cannot be
removed prior to the emergence of the subsequent ratoon
crop. To take full advantage of glyphosate as a ripener
for enhancing sugar levels at harvest and to reduce the
impact of the application on the subsequent ratoon crop,
growers should not apply glyphosate after mid-October.
In spite of the availability of a number of newly released
varieties in Louisiana since 2000, the response of many
of these varieties to the standard rate (0.21 kg ae/ha)
of glyphosate has been inconsistent and higher rates
are being applied in an attempt to elicit a response.
Richard and Dalley (2009) conducted an experiment to
investigate higher rates on a new variety that in previous
studies did not show a response at the standard rate.
Glyphosate was applied at 0.21, 0.28, and 0.35 kg ae/
ha in September 2007 to a plant-cane crop of the variety
‘L 97-128’. The two higher rates resulted in a 33-38%
increase in TRS over the non-treated control but did not
increase TRS levels over the standard rate (0.21 kg ae/
ha) at 4 WAT. At 6 WAT, the increase in TRS resulted in
a 19 to 34% increase in sugar yields for all rates based
on estimated cane yields calculated from stalk weights
and populations. At 8 WAT, however, the percent increase
in TRS was less, 9 to 10%, and there was no significant
increase in per hectare sugar yields as cane yields were
further reduced. Use of the highest rate of glyphosate
resulted in increased bleaching of the leaves, stunting,
and reduction in spring shoot numbers in the subsequent
first-ratoon crop, which translated into a 14% reduction
in first-ratoon sugar yields. Other preliminary studies on
a newly released variety,’ L 99-233’, showed similar
results of no increases in TRS with both the standard rate
of 0.21 kg ae/ha and a higher rate of 0.28 kg ae/ha.
Growers should avoid using higher rates in the hopes of
eliciting a ripening response in varieties that don’t respond
to the standard rate.
Treatments to harvest intervals (THI) of 35 to 49 days are
suggested for ripener applications of glyphosate. The
THI’s take into account the reduction in stalk elongation ,
which equates to a slight reduction in cane tonnage that
is offset by the anticipated 10 to 15% increase in sucrose
levels. Treatment to harvest interval can affect both level
of response to glyphosate (enhance per hectare sucrose
yields) and carryover injury in the subsequent ratoon
crop. Carryover injury effects have been associated
with other external stress factors such as drought. The
retention of the post-harvest blanket of straw has been
shown to reduce yields of the subsequent ratoon crop in
Louisiana, thus this stress is usually avoided by removal
of the straw soon after harvest but before the new crop
breaks winter dormancy. Often, though, there are areas
where the blanket of straw cannot be removed. Viator
et al. (2008) conducted an experiment to determine
the combined effects of post-harvest straw retention,
ripener application, and ripener treatment to harvest
intervals (THI) on yields of the treated first ratoon and
the subsequent non-treated second ratoon. Whole plots
consisted of either a non-treated control or a 0.21 kg
ae/ha application of glyphosate to first ratoon LCP 85384. Split-plots consisted of THI of 40, 50, and 60 days
for the first ratoon. Split-split plot treatments consisted of
mechanically repositioning the blanket of straw from the
row top into the wheel furrow (partial removal) compared
with complete retention (no removal) of the straw in
the second ratoon. Only the 50 day THI glyphosate
application increased sucrose yield, by 740 kg/ha when
compared to the control in the first ratoon. In the second
ratoon, there was a glyphosate by THI interaction on
sucrose yield. Harvesting at a 60 day THI in the first
ratoon reduced cane and sucrose yield of the second
ratoon by 5.4 Mg/ha and 900 kg/ha, respectively, when
compared to the averages for the 40 and 50 THI for
glyphosate-treated cane. This same treatment reduced
sucrose yields by 300 kg/ha when compared to the nontreated control in the second ratoon. Full straw retention
reduced cane and sucrose yield by 2.3 Mg/ha and
300 kg/ha compared to partial removal. It is currently
recommended that treatment to harvest intervals should
not be extended beyond 50 DAT because of possible
yield reduction in the following ratoon as well as the fact
that the continued growth in the non-treated cane along
with natural ripening negates the sugar yields associated
with the glyphosate application.
There is a continuing need to evaluate sugarcane ripeners
in order to increase the utility of the currently registered
ripener and to find additional ripeners for use by sugarcane
industries. Current research into alternative ripeners
has focused on the gibberellin inhibitor trinexapacethyl [ethyl 4-(cyclopropylhydroxymethylene)-3,5d i oxo c yc l o h ex a n e c a r b ox y l a t e ] , a n d o n t h e
ALS inhibitors imazapyr (±)-2-[4,5-dihydro4-methyl-4-(1-methylethyl)-5-oxo
-1H-imidazol-2-yl]-3-pyridinecarboxylic acid, and
nicosulfuron M2-[[[[4,6-dimethoxy-2-pyrimidinyl)
amino]carbonyl] amino]sulfonyl]-N,Ndimethyl-3pyridinecarboxamide. Richard et al. (2006) reported
an increase in TRS with LCP 85-384 treated with
glyphosate at 0.2 kg ae/ha, trinexapac-ethyl at 0.3
kg ai/ha, imazapyr at 0.1 kg ai/ha, and nicosulfuron
at 0.05 kg ai/ha when measured at 6 and 7 (WAT)
compared to a non-treated control. Sugar yields were
also increased for the trinexapac-ethyl, imazapyr, and
nicosulfuron treatments; however, there was no increase
90
Tópicos em Ecofisiologia da Cana de Açúcar
_projeto1.indb 90
13/10/10 10:22](https://image.slidesharecdn.com/ecofisiologiadacana-131108234749-phpapp02/85/Ecofisiologia-da-cana-90-320.jpg)





















Este documento é um livro sobre ecofisiologia da cana-de-açúcar publicado em 2010 pela Fundação de Estudos e Pesquisas Agrícolas e Florestais. O livro contém 12 capítulos que abordam tópicos como fenologia, modelagem de produtividade, zoneamento agrícola, interação da cana com ambientes estressantes, alterações morfofisiológicas em resposta à seca, interação entre água e nitrogênio, adubação nitrogenada, efeitos de herbicidas e reguladores de cresc



































































