Baixado 258 vezes












![Mensuração da competição
intra-específica e interespefícia
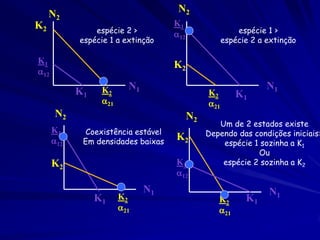
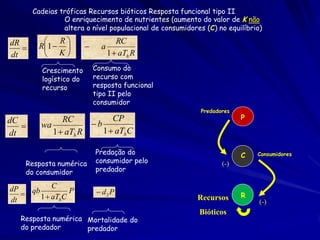
Na equação logística, o termo N/K mensura o efeito sobre
o crescimento de uma população pela adição de um membro
novo da mesma espécie (competição intra-específica).
dN1 /dt = r1N1 [(K1- N1) / K1]
E para a segunda espécie
dN2 /dt = r2N2 [(K2 - N2)/K2]
O efeito da segunda espécie sobre o crescimento da primeira
espécie. Pode ser modelado ao adicionar um segundo termo
que mede o efeito da adição de indivíduos da segunda espécie
(Competição inter-específica).
dn1/dt = r1N1(1 - N1/K1 - N2/K1)](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-17-320.jpg)
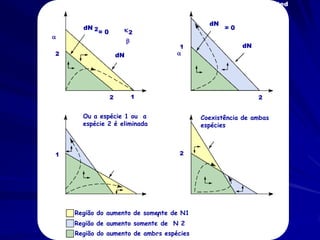
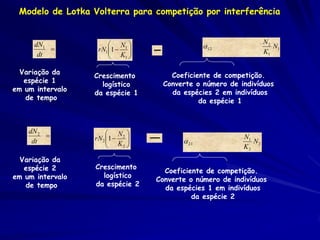
![Modelagem da Competição
Populações que competem
Podemos agora adicionar outro fator que
pode limitar a abundancia de uma espécie.
– Fator de conversão que quantifica o
efeito competitivo per capita de uma
espécie sobre outra
– Para a espécie 1:
– dN1 /dt = r1N1 [(K1 - N1 - 12 N2)/K1]](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-19-320.jpg)
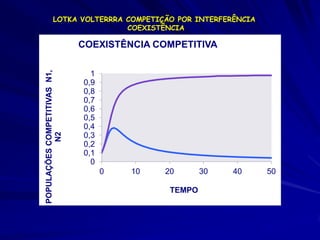
![Modelagem da Competição
Populações que competem
– Para a espécie 1;
– dN1 /dt = r1N1 [(K1 - N1 - 12 N2)/K1]
12 = efeito competitivo per capita da
espécie 2 sobre a espécie 1
21 = o efeito competitivo per capita
da espécie 1 sobre a espécie 2](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-20-320.jpg)



![Alunos e pizza: um exemplo
Numa república há 100 pedaços de pizza – o que
um aluno masculino normal faz?
Primeiro, come vários pedaços. (um aluno
masculino pode comer 10 pedaços)
Segunda, espalha a noticia as colegas
– Mas não exagere!
Terceiro, a republica chega a capacidade de
suporte com 10 alunos machos.
Assim, K=10 para alunos
machos.
dN1/dt = rN [(K – N)/K]](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-24-320.jpg)
![Alunos e pizza: um exemplo
O que acontece se um aluno macho e uma aluna
fêmea descobrem o estoque de pizza ao
mesmo tempo?
Primeiro, coma vários pedaços. (Uma aluna pode comer 5
pedaços)
Segundo, espalha a notícia a colegas
– Mas não exagere!
Terceiro, a republica chega a capacidade de suporte em
x homens e y mulheres.
Qual é a capacidade de suporte?
– Depende…
– dN1 /dt = r1N1 [(K1 – N1)/K1]
– dN12/dt = r2N2 [(K2 – N2)/K2]](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-25-320.jpg)
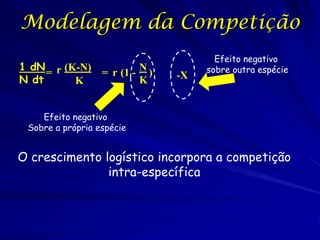
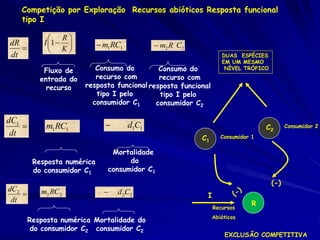
![Lotka e Volterra e Pizza
Precisamos combinar as duas equações.
Se espécies estão em competição, o número de
homens diminua com o aumenta os números de
mulheres.
De modo que:
N1 = 12 N2
Onde é o coeficiente de competição
Modelo de Lotka e Volterra: Um modelo
logístico da competição inter-específica dos
fatores intuitivos.
dN1 /dt = r1N1 [(K1 - N1 - 12 N2)/K1]](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-26-320.jpg)






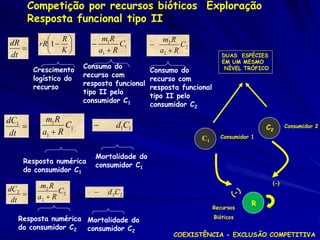
![Termina em pizza? dN1 K1 N1
r1 N1
N
dt
Na sala existe 100 pedaços de pizza. 1
– Cada aluno macho precisa 10 pedaços de pizza.
– Por isso, K1 = 10
– Mas cada aluna fêmea precisa somente 5 pedaços de
pizza.
– Por isso, K2 = 20
– Se na sala K1 existe e um aluno macho sai, quantas
fêmeas podem entrar?
Por isso, N1 = 12 N2 , onde 12 = 0.5
e, N2 = 21 N1 , onde 21 = 2
dN1 /dt = r1N1 [(K1 - N1 - 12 N2)/K1]
dN12/dt = r2N2 [(K2 – N2 - 21 N1)/K2]](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-34-320.jpg)























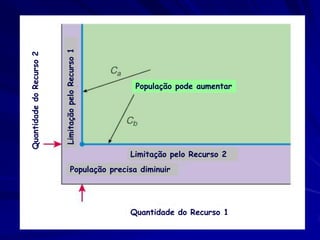

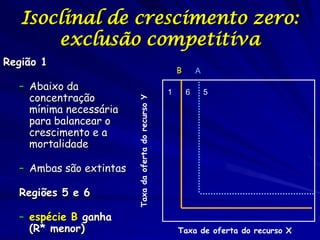
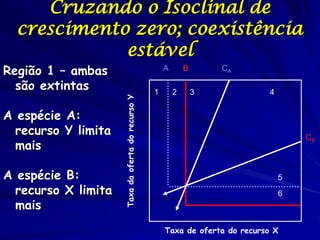
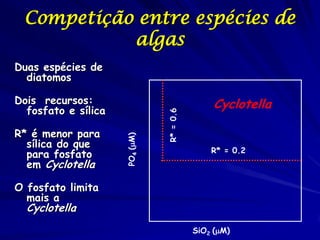

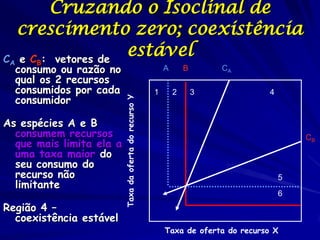
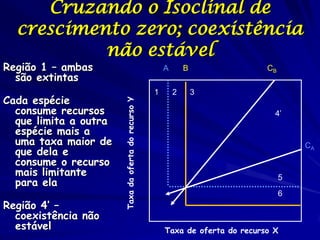
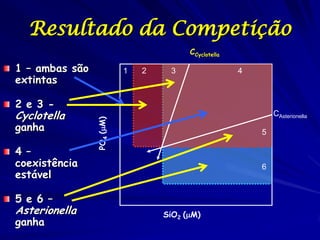
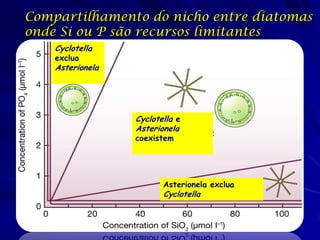
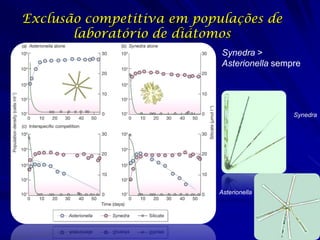
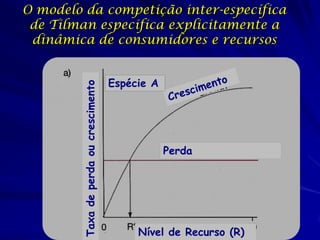
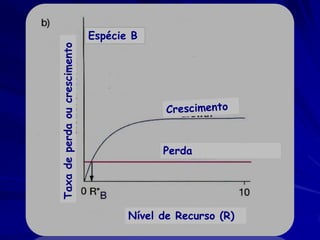
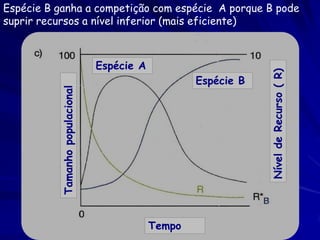
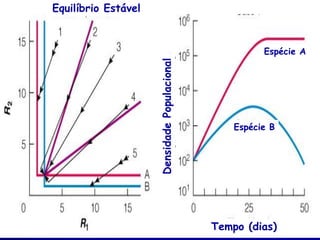
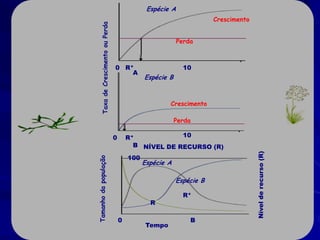

![Paradoxo do enriquecimento nas interações
competitivas (Riebesell 1974; Tilman 1982, 1988)
Tangente dos vetores de
É uma das formas pelo
consumo para A
qual as interações
competitivas também
podem resultar in a A
paradoxo do
B Tangente dos
enriquecimento vetores de
Imagine o que R2 1 2 3 consumo para B
4
acontece quando [P]
fertilizamos com N
5
Essa idéia também se 6
desenvolveu do desejo
de advertir o uso
indiscriminado de R1 [N]
enriquecimento de
recursos para Vetores de Ponto de
favorecer populações consumo Oferta de recurso
manejadas](https://image.slidesharecdn.com/competitionmodels-110516162853-phpapp02/85/Modelos-de-competicao-85-320.jpg)
















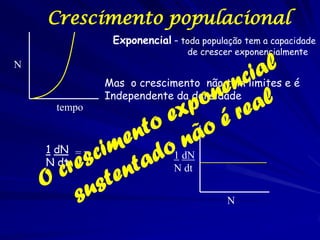
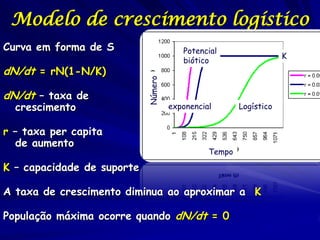
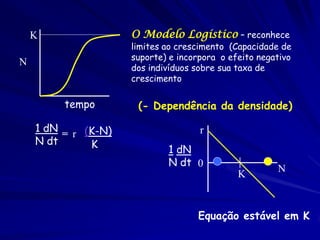
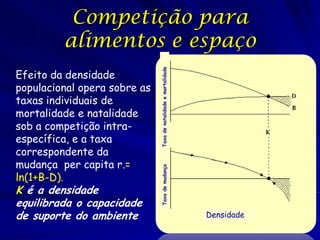
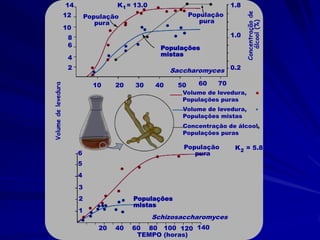
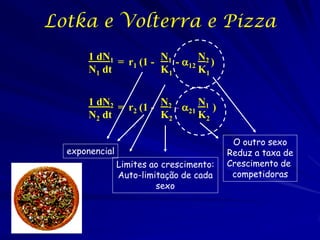
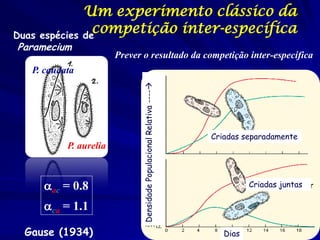
1) O documento discute modelos matemáticos de crescimento populacional, incluindo crescimento exponencial, logístico e modelos de competição entre espécies. 2) É apresentado o modelo de Lotka-Volterra para modelar a competição entre duas espécies através de equações diferenciais que descrevem as taxas de crescimento de cada população. 3) As equações incorporam termos que quantificam o efeito competitivo de cada espécie sobre a outra, permitindo modelar a competição interespecífica e prever se























































































