Baixado 161 vezes






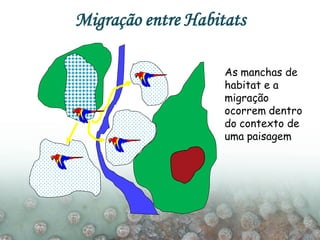




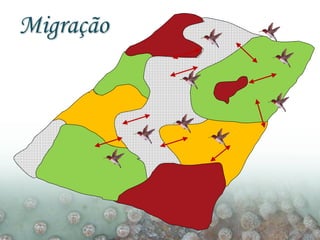


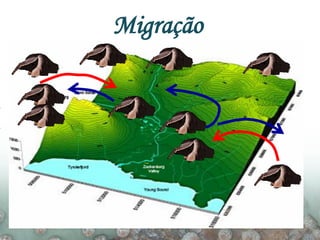




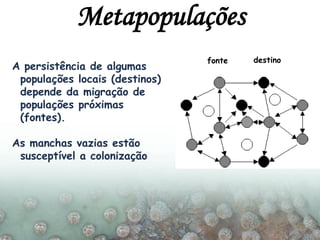







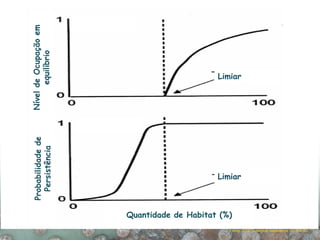














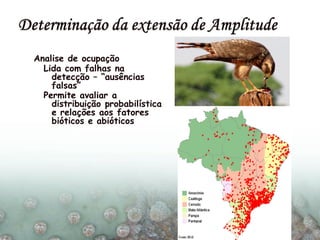








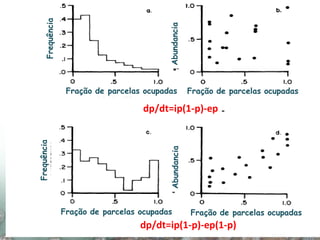








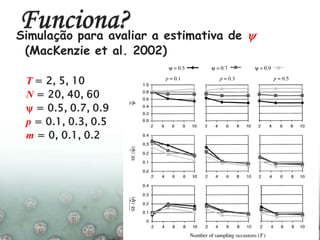










O documento discute a ecologia de populações em paisagens fragmentadas, cobrindo tópicos como processos demográficos, distribuição heterogênea de populações, migração entre habitats e o conceito de metapopulações. É introduzido o modelo de Levins para descrever a dinâmica de metapopulações e fatores que influenciam a taxa de colonização e extinção.



































































