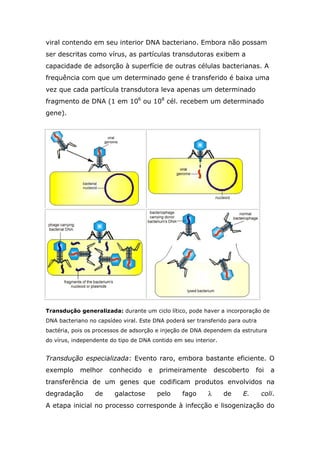
Este documento descreve quatro processos de transferência horizontal de genes entre bactérias: transformação, conjugação, transdução e conversão lisogênica. A transformação envolve a incorporação de DNA livre no meio no interior da célula. A conjugação envolve a transferência de DNA entre bactérias através do contato direto mediado por plasmídeos. A transdução envolve a transferência de DNA entre bactérias mediada por vírus. A conversão lisogênica envolve a transferência de genes de bactérias para vírus e vice-versa.