Baixado 176 vezes


















































![50 Biocontrole de doenças de plantas: uso e perspectivas
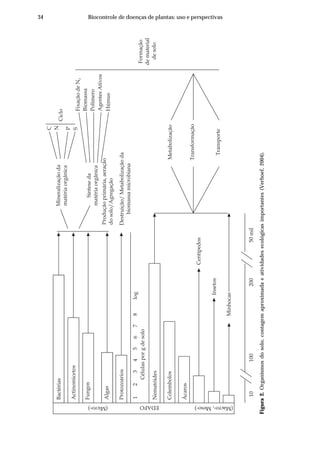
In most regions of the world soilborne plant diseases and plant-parasitic
nematodes continue to be major factors in crop losses. The primary means for
controlling soilborne plant pathogens and pests remains the application of biocidal
fumigants that kill the vast majority of organisms living in soil. However, as we
unravel the rhizosphere ecology and come to identify the multiple benefits some
organisms can confer to crop production and as the demand by society to find
reduced-risk and non-chemical options of managing soilborne diseases increase,
the use of organic amendments is again gaining attention.
Our initial objective for the use of organic amendments was as delivery
agents for microorganisms with known biological control activity. There was also
hope that these materials would stimulate the activity of not only biocontrol agents
being applied but also those of the resident microorganisms. However, we came to
the realization quite early that many such products are effective in suppressing
disease without the need for additional microorganisms. The mechanism by which
they do this we felt was worth further investigation. Summaries of this work have
been published elsewhere (Bailey & Lazarovits, 2003; Lazarovits, 2001, 2004;
Lazarovits et al., 2001, 2005). In this chapter we will focus on the two most effective
materials thus far tested, namely fish emulsion (FE) and liquid swine manure (LSM).
Both these products have unique activities and by developing an understanding of
their mode of action we expect to realize a much greater potential for their use as
fertilizers and plant protection products.
The use of fish by-products for fertilization of crops has a long history (Ceci,
1975). Fish meal, the dried protein obtained from processed fish, has been used as a
soil amendment with great success in vegetable production systems (Blatt & McRae,
1998; Gagnon & Berrouard, 1994) and was observed to reduce the populations of
plant-parasitic nematodes (Akhtar & Mahmood, 1995). FE, the concentrated fraction
of the soup left after processing fish into fish meal, is applied to crops in a diluted
form (1:100-1000) primarily as a foliar fertilizer (Aung et al., 1984). However, some
growers have reported plant health benefits and increased growth responses after
application of FE but most manufacturers do not have a clear idea as to why growers
are using their FE products.
Plant health benefits derived from using LSM are even less well documented
or understood than for FE. Our initial intent when we applied LSM to a potato field
wastodetermineifLSMincreasedtheincidenceofcommonscab,asthevastmajority
of literature indicated that manure applications would have this effect. However,
we discovered that a single application of LSM to a potato field having a soil pH of
5 resulted in a significant reduction of common scab [Streptomyces scabies (Thaxter)
Lambert & Loria = Streptomyces. scabiei (Trüper & dè Clari, 1997)] and verticillium
wilt [Verticillium dahliae Kleb.] for up to 3 years (Conn & Lazarovits, 1999). Disease
reduction with the same LSM, however, was not observed at a second site where the
soil pH was neutral (Conn & Lazarovits, 1999). At that time we were not able to
explain this anomaly. To understand what mechanisms may be acting we brought
the soil and the LSM back to the laboratory and studied the effects using microcosm
systems. With both and LSM and FE, small scale laboratory model systems proved
to be a critical tool for identifying how these materials impacted plant pathogen
populations and hence disease severity.](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-51-320.jpg)
































































































































![198 Biocontrole de doenças de plantas: uso e perspectivas
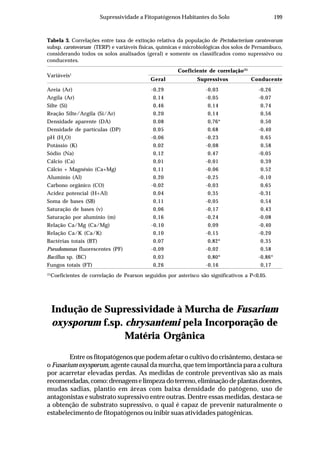
Um aspecto a ser considerado na dificuldade da caracterização dos possíveis
mecanismos envolvidos na supressividade dos solos de Pernambuco à população
do Pcc127Rif
é a metodologia, uma vez que foram utilizados indicadores tradicionais
de supressividade (Hornby, 1983; Chellemi & Porter, 2001). De acordo com Van
Bruggen&Semenov(1999),osresultadosobtidosutilizandoindicadorestradicionais
são difíceis de interpretar, motivo pelo qual sugeriram um procedimento alternativo,
baseado na mensuração de respostas biológicas a distúrbios ou estresse, assumindo
que um solo sadio é estável, com resistência ao estresse. A resposta ao estresse em
termos de amplitude e resistência da comunidade microbiana poderia ser um
indicador universal para supressão à doença melhor que qualquer outro fator físico,
químico ou biológico mensurado somente uma vez a longos intervalos de tempo.
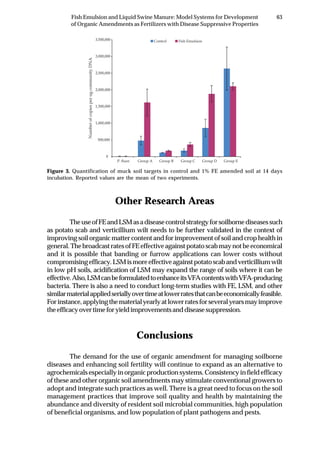
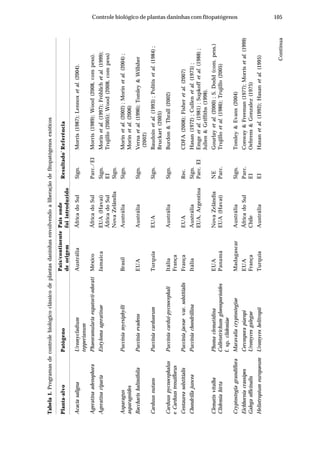
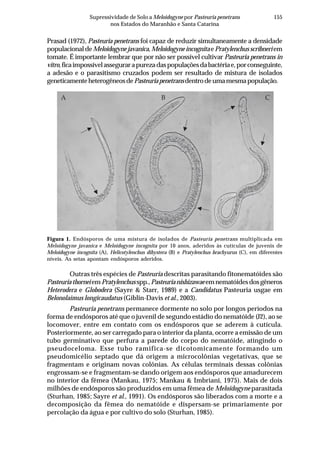
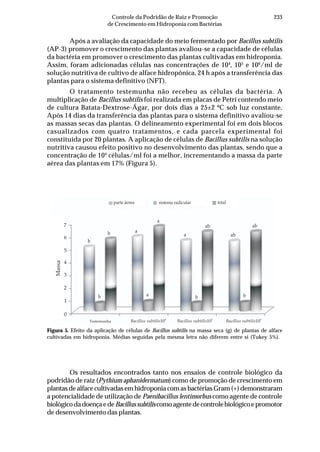
Tabela 2. Influência de solos do estado de Pernambuco na taxa de extinção relativa da população
(TERP) de Pectobacterium carotovorum subsp. carotovorum
Código do solo Local (município) Cobertura(1)
Textura TERP2
CF-1 Camocim de São Félix couve-chinesa Franco arenoso 0,6327 a3
SA-1 Sairé milho Franco argilo arenoso 0,5439 b
BG-1 Barra de Guabiraba sem plantio Franco argilo arenoso 0,4011 c
CO-1 Condado cana-de-açúcar Argila 0,3682 c
GR-1 Gravatá couve-chinesa Franco arenoso 0,3357 c
GR-2 Gravatá pimentão Franco arenoso 0,3145 c
CF-2 Camocim de São Félix couve-chinesa Franco argilo arenoso 0,2801 d
AL-1 Abreu e Lima cana-de-açúcar Areia franca 0,2315 d
CG-1 Chã Grande brócoli Franco arenoso 0,2301 d
CF-3 Camocim de São Félix couve-chinesa Franco argilo arenoso 0,2004 d
BO-1 Bonito mata Argila 0,2003 d
SJ-1 São Joaquim do Monte pimentão Franco argilo arenoso 0,1992 d
CF-4 Camocim de São Félix pimentão Franco argilo arenoso 0,1722 d
SJ-2 São Joaquim do Monte pimentão Franco arenoso 0,1667 d
CG-2 Chã Grande pimentão Franco arenoso 0,1519 e
CF-5 Camocim de São Félix couve-chinesa Franco argilo arenoso 0,1483 e
GR-3 Gravatá couve-chinesa Franco argilo arenoso 0,1397 e
CF-6 Camocim de São Félix couve-chinesa Franco argilo arenoso 0,1238 e
AN-1 Aliança cana-de-açúcar Franco arenoso 0,1121 e
IG-1 Igarassu inhame Franco arenoso 0,0914 f
CO-2 Condado cana-de-açúcar Franco arenoso 0,0806 f
CF-7 Camocim de São Félix couve-chinesa Franco argilo arenoso 0,0799 f
CF-8 Camocim de São Félix couve-chinesa Franco arenoso 0,0718 f
CF-9 Camocim de São Félix couve-chinesa Franco arenoso 0,0547 f
C.V. (%) 10,34
(1)
Tipo de cobertura do solo na época da coleta. 2
Taxa de extinção relativa da população de
Pectobacterium carotovorum subsp. carotovorum no solo [log UFC/dia], calculada conforme Kocks
et al. (1998). 3
Média de quatro repetições. Médias seguidas pela mesma letra não diferem entre si
(Scott-Knott P=0,05).](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-196-320.jpg)



![206 Biocontrole de doenças de plantas: uso e perspectivas
Hornby, D. Suppressive soils. Annual Review of Phytopathology 21: 65-85. 1983.
Hornby, D. (Ed.) Biological Control of Soil-borne Plant Pathogens. Wallingford. CAB International.
1990.
Huber, D.M. & Schneider, R.W. The description and occurrence of suppressive soils. In: Schneider,
R.W. (Eds.) Suppressive Soils and Plant Disease. St. Paul. The Americam Phytopathological
Society. 1982. pp. 1-7.
Hyakumachi, M. Suppression and prevention in Rhizoctonia disease: mechanisms involved in
disease decline. Palestras, 5
º
Simpósio de Controle Biológico, Foz do Iguaçu, PR. 1996. pp.
140-149.
Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Matteile, T. & Steinberg, C. Soil
health through soil disease suppression: Which strategy from descriptors to indicators? Soil
Biology & Biochemistry 39: 1-23. 2007.
Jones, J. P. Fusarium wilt. In: Jones, J. B.; Jones J. P.; Stall, R. E. & Zitter, T. A. (Eds.) Compendium
of Tomato Diseases. St. Paul. APS Press. 1991. p. 15.
Kikumoto, T. Ecological aspects of soft rot bacteria. Report of the Institute for Agricultural
Research (Tohoku University) 31: 19-41. 1980.
Kikumoto, T. Ecology and biocontrol of soft rot of Chinese cabbage. Journal of General Plant
Pathology 66: 275-277. 2000.
Kim, B-S. & Yeoung, Y.R. Suppression of bacterial rot on Chinese cabbage by calcium fertilizer
treatment. Research in Plant Disease 10: 82-85. 2004.
Kocks, C.G.; Ruissem, M.A.; Zadoks, J.C. & Duijkers, M.G. Survival and extinction of Xanthomonas
campestris pv. campestris in soil. European Journal of Plant Pathology 104: 911-923. 1998.
Komada, H. Development of a selective medium for quantitative isolation of Fusarium oxysporum
from natural soil. Review of Plant Protection Research 8: 114-125. 1975.
Kurozawa, C. & Pavan, M. A. Doenças das cucurbitáceas. (abóbora, abobrinha, chuchu, melancia,
melão, moranga, pepino). In: Kimati, H.; Amorim, L.; Bergamin Filho, O. A.; Camargo, L. E.
A. & Rezende, J. A. M. Manual de Fitopatologia. Vol. 2. 3ª. Ed. São Paulo. Ceres. 1997a. pp.
325-337.
Kurozawa, C. & Pavan, M. A. Doenças do tomateiro. In: Kimati, H.; Amorim, L.; Bergamin Filho,
O. A.; Camargo, L. E. A. & Rezende, J. A. M. Manual de Fitopatologia. Vol. 2. 3ª. Ed. São
Paulo. Ceres. 1997b. pp. 690-719.
Larkin, R.P.; Hopkins, D.L. & Martin, F.N. Suppression of Fusarium wilt of watermelon by
nonpathogenic Fusarium oxysporum and other microorganisms recovered from a disease-
suppressive soil. Phytopathology 86: 812-819. 1996.
Lartey, R.T.; Curl, E.A.; Peterson, C.M. & Harper, J.D. Mycophagous grazing and food preference
of Proiisotoma minuta (Collembola: Isotomidae) and Onychiurus encarpatus (Collembola:
Onychiuridae). Environmental Entomology 18: 334-337. 1989.
Laterrot H.; Blancard, D. & Couteaudier Y. Les Fusarioses de la tomate. P. H. M. Revue Horticole.
1988. 288 p.
Lazarovits, G. Managing soilborne plant diseases through selective soil disinfestation by a knowledge
based application of soil amendments. Phytoparasitica 32: 427-431. 2004.
Lazarovits, G.; Tenuta, M. & Conn, K.L. Organic amendments as a disease control strategy for
soilborne diseases of high-value agricultural crops. Australasian Plant Pathology 30: 111-
117. 2001.
Lazarovits, G.; Conn, K.L. & Abbasi, P.A. Understanding the mode of action of organic soil
amendments provides the way for improved management of soilborne plant pathogens. In:
Vanachter, A. (Ed.) Proceedings of the Sixth International Symposium on Chemical an Non-
Chemical Soil and Substrate Desisfestation. [S.n., 2005]. pp. 215-222.
Lazarovits, G.; Abassi, P. & Conn, K. Managing soil agro-ecosystems for environmental and plant
health: back to the future. Summa Phytopathologica 32: 153-156. 2006.
Lopes, C. A.; Reis, A. & Àvial, A. C. Doenças do tomateiro para mesa causadas por fungos,
bactérias e vírus. Informe Agropecuário 24: 66-78. 2003.
Lumsden, R.D. Development of Gliocladium virens for biological control of Pythium and Rhizoctonia
damping-off diseases of seedlings. Proceedings, 13
rd
International Plant Protection Congress,
The Hague, The Netherlands. 1995. pp. 385-388
Luz, W. C. Rizobactérias promotoras de crescimento de plantas e de bioproteção. Revisão Anual
de Patologia de Plantas 4: 1-47, 1996.](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-204-320.jpg)













































![266 Biocontrole de doenças de plantas: uso e perspectivas
Silva, L.F. & Leite, J.O. Caracterização preliminar dos agrossistemas das regiões cacaueiras da
Bahia e Espírito Santo. In: CEPLAC (Ed.) Boletim Técnico 156. Ilhéus BA. 1970. pp.15.
Silva Neto, P.J. Sistema de produção de cacau para a Amazônia Brasileira. Belém. CEPLAC.
2001.
Silva, S.D.V.M.; Gramacho, K.P. & Almeida, O.C. Solanum paniculatum hospedeiro de Crinipellis
perniciosa na região sul da Bahia. Agrotrópica 4: 17-20. 1992.
Silva, S.D.V.M.; Luz, E.D.M.N.; Almeida, O.C.; Gramacho, K.P. & Bezerra, J.L. Redescrição da
sintomatologia causada por Crinipellis perniciosa em cacaueiro. Agrotrópica 14: 1-24. 2002.
Singer, R. A monographic study of the genera Crinipellis and Chaetocalathus. Lilloa 8: 441–534.
1942.
Souza, J.T.; Pomella, A.W.V.; Bowers, J.H.; Pirovani, C.P.; Loguercio, L.L. & Hebbar, K.P. Genetic
and biological diversity of Trichoderma stromaticum, a mycoparasite of the cacao witches’-
broom pathogen. Phytopathology 96: 61-67. 2006.
Stadinik, M.J. & Talamini, V. Legislação e uso de produtos fitossanitários naturais em países do
cone sul. In: Stadinik, M.J. & Talamini, V. (Eds.) Manejo ecológico de doenças de plantas.
Florianópolis. CCA/UFSC. 2004. pp. 63-82.
Stahel, G. Marasmius perniciosus nov. spec., the cause of Krulloten disease of cocoa in Suriname.
Paramarimbo. In: Dept. Van Den Landbouw (Ed.) Bulletin 33. Suriname. 1915. pp. 1-27.
Thorold, C.A. Diseases of Cocoa. Oxford. Clarendon Press. 1975.
Tovar, G. La escoba de bruja del cacao [Crinipellis perniciosa (Stahel) Singer]: descripción de sintomas
de la enfermedad. Agronomia Colombiana. 8: 227-239. 1991.
Tronsmo, A. & Hjeljord, L.G. Biological control with Trichoderma species. In: Boland, G.J. &
Kuykendall, L.D. (Eds.) Plant-microbe interactions and biological control. New York. Marcel
Dekker. 1998. pp. 111-126.
Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 22: 837-845.
1932.
Wood, G.R.A. & Lass, R.A. Cocoa. 4
th
Ed. London. Longman. 1985.](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-262-320.jpg)























![293Controle Biológico de Doenças de Flores e Frutos Jovens de Citros
limoeiro ‘Cravo’, com 12 anos de idade, com espaçamento de 7 x 3,5 m,
localizado no município de Rincão, SP. A multiplicação dos isolados de
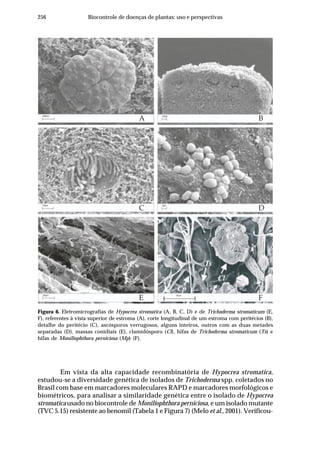
Bacillus subtilis (ACBs 69, 72, 77 e AP-3) foi em meio à base do resíduo da
fermentação glutâmica do melaço (Aminofértil®
) a 5%, durante 72 h (Bettiol
& Astiarraga, 1998). Na safra 2002/2003 os estudos foram conduzidos em
pomar de laranjeira ‘Natal’ enxertada sobre limoeiro ‘Cravo’, com oito anos
de idade, localizado no município de Santa Rita do Passa Quatro, SP, em
área de notório histórico da doença. Além dos isolados de Bacillus subtilis
(AP-3, ACB 69, 72 e 77) testaram-se os isolados de Trichoderma, Trichoderma
viride (ACB-14) e Trichoderma sp. (ACB-40) que inibiram o patógeno in vitro.
A multiplicação de Bacillus subtilis foi em meio à base de farelo de algodão
(Bettiol et al., 2005). A multiplicação de Trichoderma foi em meio à base de
batata-dextrose. Os caldos fermentados por Bacillus subtilis ou Trichoderma
foram diluídos a 10% e comparados com o tratamento padrão [cobre metálico
(100 g de oxicloreto de cobre/100 l de água) e carbendazim + mancozeb (25 g
de i.a. + 160 g de i.a./100 l de água, respectivamente, acrescidos de óleo
mineral a 0,5% (Fundecitrus, 1998)]. Na safra 2001/2002 avaliou-se, também,
a mistura dos isolados de Bacillus subtilis (ACBs 69+72+77+AP-3) e um
biofertilizante na concentração de 10% (Kupper et al., 2006). A primeira
pulverização foi realizada em 15/10/2001 (após a queda de 3/4 das pétalas).
As pulverizações com Bacillus subtilis foram repetidas 28, 56, 84, 112 e 140
dias após a primeira. Para o tratamento padrão foram realizadas duas
aplicações com oxicloreto de cobre, na data da primeira pulverização e 28
dias após e duas com a mistura carbendazim + mancozeb aos 84 e 140 dias
após a primeira pulverização. A mistura de isolados de Bacillus foi preparada
utilizando-se proporções iguais dos isolados. Na safra 2002/2003, no
primeiro ensaio, os antagonistas e os fungicidas foram aplicados de acordo
com o esquema descrito para a safra 2001/2002, cuja primeira aplicação foi
realizada em 08/10/2002. No outro ensaio, as pulverizações com os agentes
de biocontrole foram quinzenais, enquanto os fungicidas foram aplicados de
forma semelhante à safra 2001/2002, sendo a primeira pulverização em 23/
10/2002.
Fungicidas e antagonistas foram aplicados com pulverizador tratorizado,
dotado de duas pistolas. O volume de calda aplicado foi calibrado para atingir o
ponto de escorrimento, sendo utilizados 100 l de calda/tratamento. As
concentrações de células nos caldos fermentados com Bacillus foi de 108
UFC/ml
na safra 2001/2002 e de 109
UFC/ml e 107
esporos/ml para Bacillus subtilis e
Trichoderma, respectivamente, na safra 2002/2003. As avaliações foram realizadas
por ocasião da colheita, em 50 frutos coletados ao acaso da planta central de cada
parcela, com o auxílio de uma escala de notas de severidade da mancha preta dos
frutos cítricos, sendo: 0 = ausência de sintomas; 1 = 0,9% até 6 = 17,3% de área
coberta com sintomas (Fagan, 1999). Posteriormente, determinou-se o índice de
doença [ID = Σ ((fv) x 100)/n), onde f = número de frutos com determinado grau de
infecção; v = grau de infecção e n = número total de frutos amostrados]. O
delineamento experimental foi o de blocos ao acaso, com quatro e cinco repetições
nos ensaios da safra 2001/2002 e 2002/2003, respectivamente. Cada parcela
experimental foi constituída por três plantas.](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-287-320.jpg)









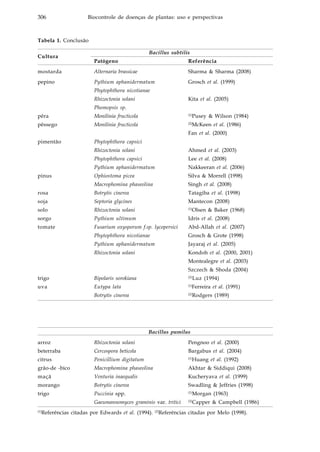


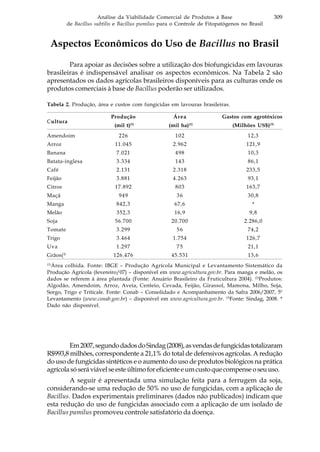








![318 Biocontrole de doenças de plantas: uso e perspectivas
Existem diversos fungicidas registrados para o controle da maioria das
doenças presentes na cultura do arroz, porém sua utilização eleva os custos de
produção, polui o ambiente, ameaça a saúde do agricultor, além de pressionar a
seleção de populações do patógeno resistentes a estes compostos químicos. Aliado
a isso, há ainda a pressão da sociedade por alimentos sem resíduos de agrotóxicos,
o que torna urgente a busca por alternativas no controle de doenças em cultivos de
arroz.
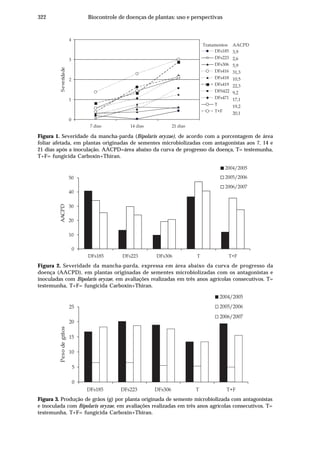
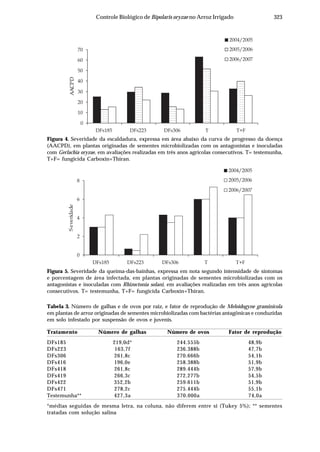
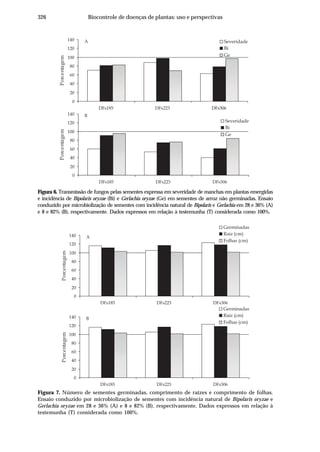
Dentre as principais doenças no arroz destaca-se a brusone, causada por
Pyricularia grisea (Cooke) Sacc. [teleomorfo: Magnaporthe grisea (Herbert) Barr]. Seu
ataque, nos estádios de plântula ou no perfilhamento, assim como infecções severas
nas panículas, pode comprometer a produção (Ou, 1985). A ocorrência desta doença
está associada às condições de temperatura amena, alta umidade relativa e grande
pressão de inóculo na lavoura, sendo particularmente importante em cultivos
irrigados no Centro-Oeste do Brasil, pois além de ser favorecida pelas condições
climáticas, o manejo da água de irrigação geralmente é deficiente.
Danos cada vez maiores são relacionados à ocorrência da queima-das-
bainhas, incitada por Rhizoctonia solani Kuhn [teleomorfo: Thanatephorus cucumeris
(Frank) Donk]. Esta doença causa redução da produtividade, devido,
principalmente, à esterilidade de espiguetas e ao acamamento das plantas. O fungo
é capaz de sobreviver no solo como estruturas de resistência (escleródios) que
posteriormente são disseminadas pela água de irrigação, infectando folhas e bainhas,
atingindo até a base da haste. O cultivo do arroz sucedido por hospedeiros deste
fungo, como a soja ou pastagens, leva à continuidade do ciclo do patógeno e ao
aumento de inóculo. Estas duas peculiaridades fazem com que os danos causados
pela queima-das-bainhas sejam crescentes a cada ano.
A escaldadura causada por Gerlachia oryzae (Hashioka & Yokogi) W. Gams
[(teleomorfo: Monographella albescens (Thümen) Parkinson, Sivanesan & C. Booth)
(sinonímia: Rhynchosporium oryzae, Microdochium oryzae)] ocorre em níveis
considerados altos em todas as regiões produtoras de arroz no Brasil, embora ainda
não existam estimativas relativas aos prejuízos causados por essa enfermidade. Os
principais danos referem-se à desuniformidade das lavouras (Prabhu & Bedendo,
1990) e à diminuição da área fotossintetizante (Webster & Grunnel, 1992).
A ocorrência de mancha-parda, causada por Bipolaris oryzae (Breda de Haan)
Shoem[teleomorfo:Cochliobolusmiyabeanus(Ito&Kuribayashi)(sinonímia:Drechslera
oryzae, Helminthosporium oryzae)], está crescendo devido ao uso de cultivares com
maior grau de suscetibilidade, tendo assumido posição de doença economicamente
importante, surgindo como principal doença do arroz em algumas regiões. Os seus
danos vão desde redução no percentual de germinação, no estabelecimento de
plantas e na área fotossintetizante, até manchas nos grãos que, além de depreciarem
o produto, geram perdas durante o beneficiamento.
Os prejuízos causados por doenças em arroz ocorrem tanto em zonas
temperadas quanto tropicais, principalmente onde o seu cultivo utiliza grandes
quantidades de fertilizantes e cultivares com alto índice de perfilhamento. Devido
à limitação do emprego de cultivares resistentes, bem como aos aspectos
econômicos e ambientais associados ao controle químico, pesquisas estão sendo
realizadas na busca de tecnologias alternativas. Diante desse cenário, o controle](https://image.slidesharecdn.com/livrobiocontrole-130513195442-phpapp02/85/Livrobiocontrole-311-320.jpg)

















O documento discute a história e situação atual do controle biológico de doenças de plantas no Brasil. Em três frases, resume: 1) O uso intensivo de agrotóxicos tem causado problemas ambientais e levado à resistência de patógenos, mas o controle biológico é uma alternativa promissora para reduzir o uso desses produtos; 2) Preocupações com impactos da agricultura fizeram emergir mercados de alimentos produzidos sem agrotóxicos, aumentando o interesse pelo controle bioló




























![[EN] Aastra - Vertical Market - Industry](https://cdn.slidesharecdn.com/ss_thumbnails/vertical-marketindustry07-2010-110407025956-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)

























































