Transferir como PDF, PPTX


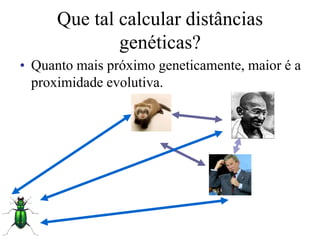



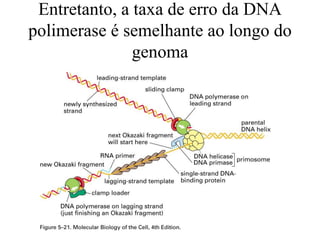

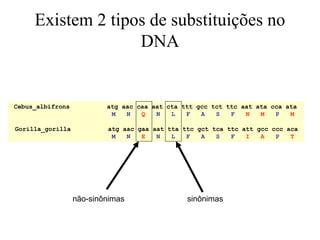
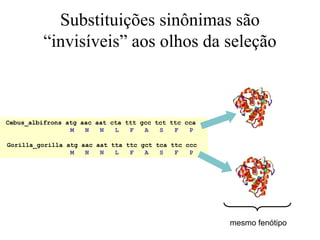
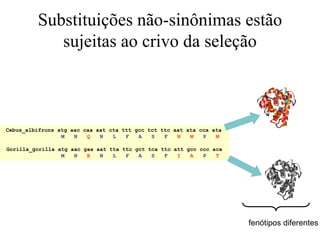
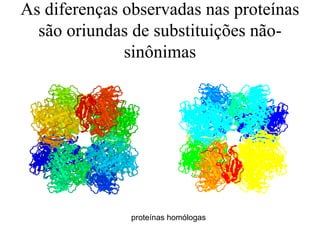


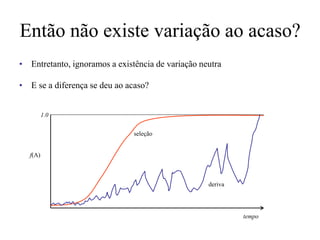



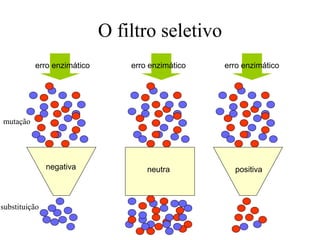



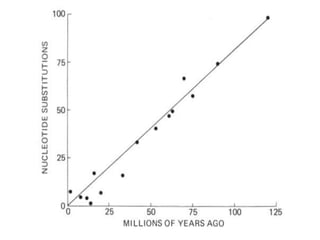


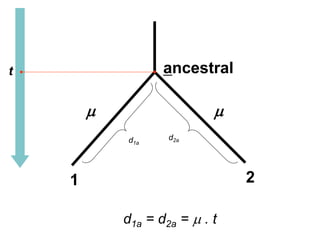
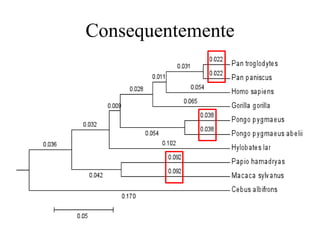


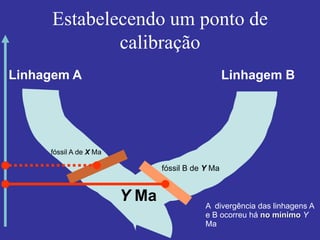

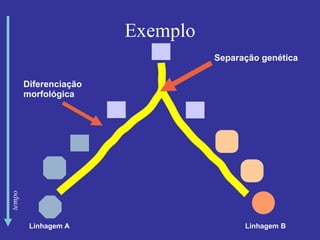

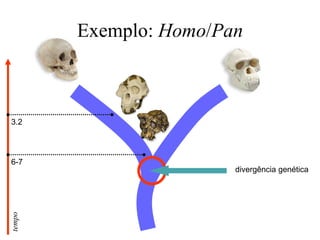
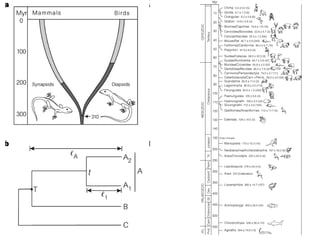

O documento discute métodos para estimar distâncias genéticas e relacionamentos evolutivos entre espécies. Ele apresenta como calcular taxas evolutivas entre genes homólogos e como usar essas taxas para estimar quando linhagens divergiram, estabelecendo um "relógio molecular". O documento também discute como distinguir variação neutra de adaptações moleculares.























































































