Baixado 499 vezes


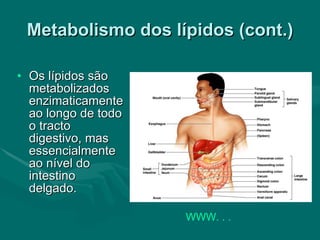

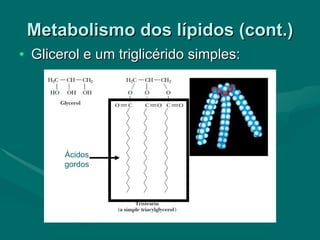
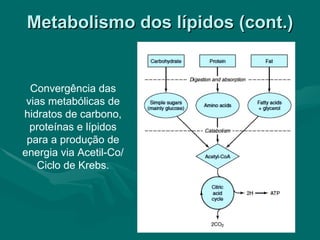
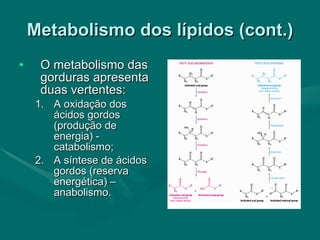
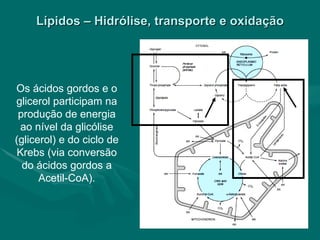

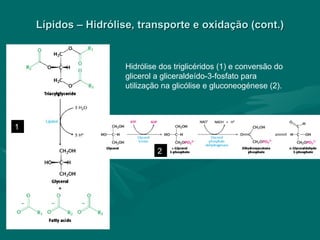

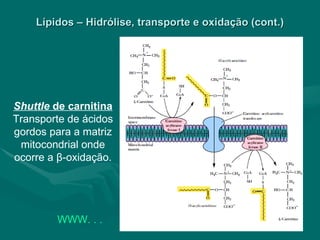

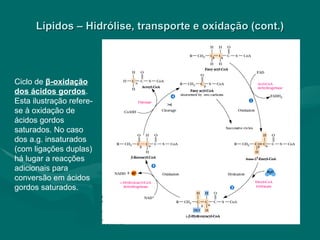



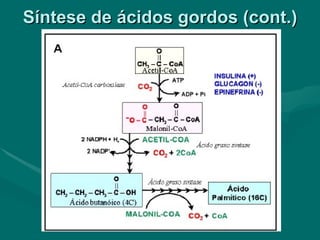
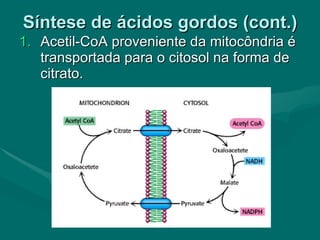
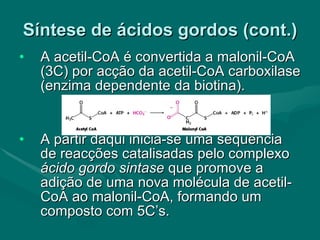




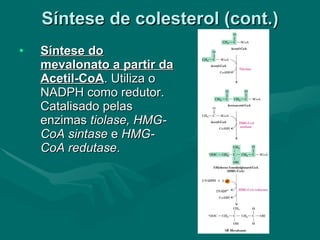

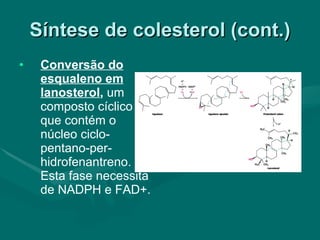
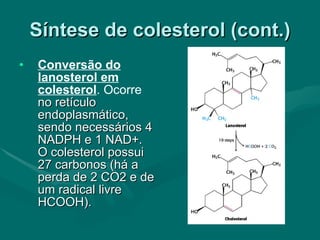


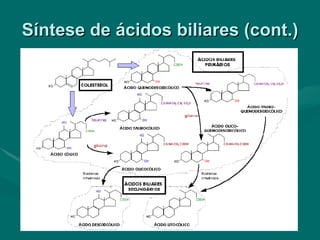

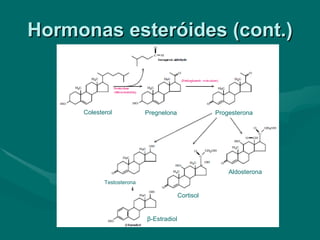


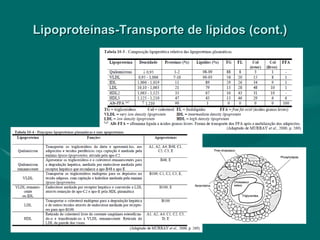


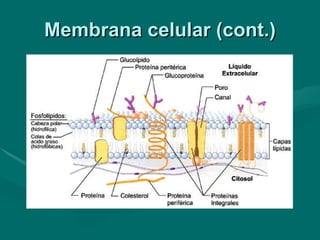



O documento discute o metabolismo dos lípidos no corpo humano. Os principais pontos são: (1) Os lípidos são uma importante fonte de energia e podem ser armazenados como gordura; (2) Os lípidos são metabolizados através de processos catabólicos que liberam energia e processos anabólicos que sintetizam novos lípidos; (3) Os principais processos envolvidos no metabolismo dos lípidos incluem a lipólise, β-oxidação e síntese de ácidos grax
































































