Baixado 238 vezes




![6 / GEOFFREY M. COOPER • ROBERT E. HAUSMAN
formam, por polimerização espontânea, as macromoléculas. Por exemplo, o aqueci-
mento de misturas secas de aminoácidos resulta na polimerização para formar poli-
peptídeos. Contudo, a característica essencial da macromolécula a partir da qual a
vida evoluiu deve ter sido a capacidade de auto-replicação. Somente uma macromo-
lécula capaz de controlar a síntese de novas cópias de si própria poderia ser capaz de
reprodução e posterior evolução.
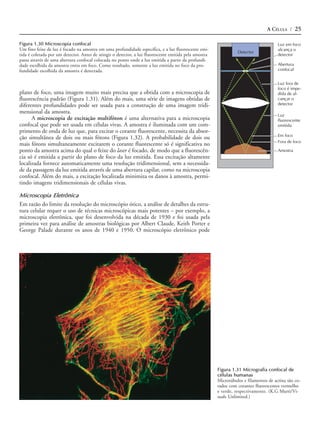
Dos dois tipos principais de macromoléculas informativas presentes atualmen-
te (ácidos nucléicos e proteínas), somente os ácidos nucléicos são capazes de contro-
lar sua auto-replicação. Os ácidos nucléicos podem servir de moldes para sua própria
síntese como resultado do pareamento específico de bases entre nucleotídeos com-
plementares (Figura 1.3). A etapa essencial no entendimento da evolução molecular
foi alcançada no início da década de 1980, quando foi descoberto nos laboratórios
de Sid Altman e Tom Cech que o RNA é capaz de catalisar várias reações químicas,
incluindo a polimerização de nucleotídeos. Estudos mais avançados ampliaram as
atividades catalíticas conhecidas do RNA, incluindo a descrição de moléculas de
RNA que controlam a síntese de uma nova fita de RNA a partir de um RNA-molde.
O RNA é, assim, tanto capaz de servir como molde quanto capaz de catalisar sua
própria replicação. Conseqüentemente, em geral é aceito que o RNA tenha sido o
sistema genético inicial, e supõe-se que a fase inicial da evolução química tenha sido
baseada nas moléculas de RNA auto-replicativas – um período da evolução conheci-
do como mundo de RNA. Então, interações ordenadas entre RNA e aminoácidos
evoluíram para o código genético atual, e o DNA finalmente substituiu o RNA
como material genético.
Presume-se que a primeira célula tenha originado-se da inclusão de RNAs auto-
replicativos em uma membrana composta de fosfolipídeos (Figura 1.4). Como dis-
cutido em detalhes no próximo capítulo, os fosfolipídeos são os componentes bási-
cos de todas as membranas biológicas atuais, incluindo as membranas plasmáticas de
células procarióticas e eucarióticas. A característica-chave dos fosfolipídeos que for-
mam as membranas é que eles são moléculas anfipáticas, significando que uma por-
ção da molécula é solúvel em água e a outra porção é insolúvel. Os fosfolipídeos têm
longas caudas de hidrocarbonetos insolúveis em água (hidrofóbica) ligadas a uma
cabeça com grupos fosfato solúvel em água (hidrofílica). Quando colocados na água,
os fosfolipídeos agregam-se espontaneamente em uma bicamada, com suas cabeças
com grupos fosfato na porção exterior em contato com a água e suas caudas de
hidrocarbonetos no interior em contato umas com as outras. Tais bicamadas de fos-
folipídeos formam uma barreira estável entre dois compartimentos aquosos – por
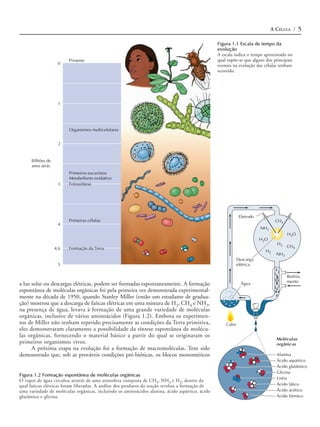
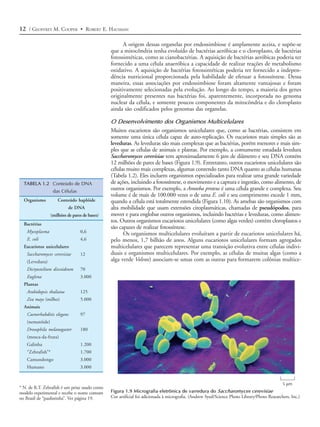
Figura 1.3 Auto-replicação do RNA exemplo, separando o interior de uma célula do meio externo.
O pareamento complementar entre nucleo-
tídeos (adenina [A] com uracil [U] e guani-
A inclusão do RNA auto-replicativo e de moléculas associadas em uma mem-
na [G] com citosina [C]) permite que uma brana de fosfolipídeos poderia tê-los mantido, assim, como uma unidade, capaz de
fita de RNA sirva como molde para a sínte- auto-replicação e posterior evolução. A síntese de proteína controlada por RNA já
se de uma nova fita com a seqüência com-
plementar.
C
C C G C G C G C G G
G
G G C G C G C G C C
A A U A U A U A U U
A
G G C G C G C G C C
A A A U A U A U
U
U U
U U A U A U A U A
U U U A U A U A U A
A
G G G C G C G C G C
C
A A U A U A U A U A U
C C C G C G C C G
G G](https://image.slidesharecdn.com/introduoabilogiacelular-130224145236-phpapp01/85/Introducao-a-biologia-celular-Leitura-complementar-5-320.jpg)

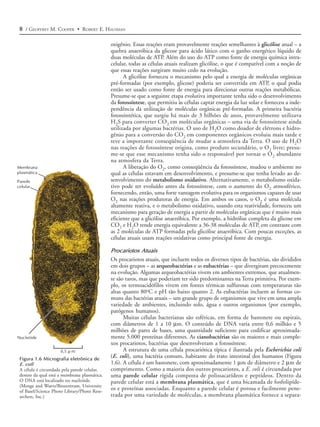


![14 / GEOFFREY M. COOPER • ROBERT E. HAUSMAN
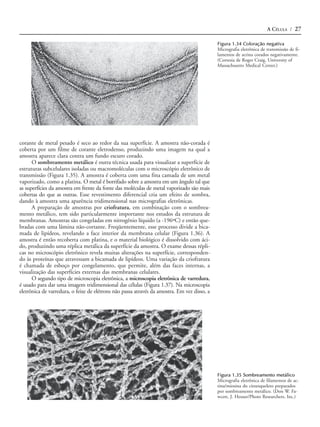
e músculo (Figura 1.13). As células epiteliais formam camadas que cobrem a super-
fície do corpo e recobrem os órgãos internos. Há muitos tipos diferentes de células
epiteliais, cada um especializado para uma função específica, incluindo proteção (a
pele), absorção (por exemplo, as células da mucosa do intestino delgado) e secreção
(por exemplo, as células da glândula salivar). O tecido conectivo inclui ossos, cartila-
gens e tecido adiposo, cada um formado por diferentes tipos de células (respectiva-
mente, osteoblastos, condrócitos e adipócitos). O tecido conectivo frouxo, que intercala
(A)i Boca (A)ii Ducto biliar (A)iii Intestino
(B)
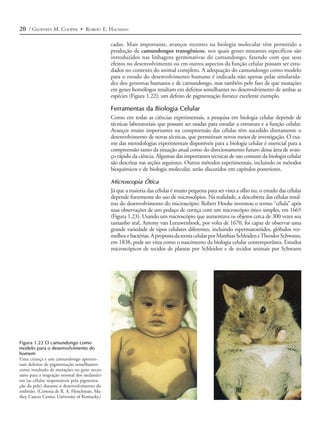
Figura 1.13 Micrografias óticas de
células animais representativas
(A) Células do epitélio da boca (uma grossa
camada multicelular), do ducto biliar e do
intestino. (B) Fibroblastos são células do te-
cido conectivo, caracterizados por sua for-
ma alongada. (C) Eritrócitos, granulócitos,
linfócitos e monócitos no sangue humano.
([A]i e [A]ii, G. W. Willis/Biological Photo
Service; [A]iii, Biophoto Associates/Photo
Researchers, Inc.; B, Don W. Fawcett/Visu-
als Unlimited; C. G. W. Willis/Biological
Photo Service.)
(C) Eritrócito Linfócito Monócito
Granulócito](https://image.slidesharecdn.com/introduoabilogiacelular-130224145236-phpapp01/85/Introducao-a-biologia-celular-Leitura-complementar-13-320.jpg)















![A CÉLULA / 39
8. Que vantagem o uso da proteína permitem a separação, em um gradien- apropriado para a análise dos movimen-
verde fluorescente (GFP) tem sobre o te de sacarose, pela centrifugação por tos de células animais?
uso de anticorpos marcados com fluo- velocidade e pela centrifugação de equi-
rescência, no estudo da localização e líbrio. 11. Por que a capacidade de cultivar célu-
do movimento de uma proteína nas las-tronco embrionárias é importante?
células? 10. As leveduras têm sido usadas como
um modelo para o estudo de muitos 12. Qual a diferença entre culturas ce-
9. Identifique as diferentes característi- aspectos da biologia das células eucarió- lulares primárias e linhagem de células
cas das propriedades das organelas que ticas. Por que elas não são um modelo imortalizadas?
Referências e Leituras Adicionais Earth before 3,800 million years ago. Na- Neidhardt, F. C., R. Curtiss III., J. L. In-
ture 384:55-59. [P] graham, E. C. C. Lin, K. B. Low Jr., B.
Orgel, L. E. 1998. The origin of life – a review Magasanik, W. Reznikoff, M. Riley, M.
A Origem e a Evolução das Células of facts and speculations. Trends Biochem. Sci. Schaechter and [H. E. Umbarger (eds.).
23: 491-495. [R] 1996. Escherichia coli and Salmonella: Ce-
Andersson, S. G. E., A. Zomorodipour, J. O. Pace, N. R.1997. A molecular view of microbi- llular and Molecular Biology. 2nd. ed. Wa-
Andersson, T. Sicheritz-Ponten, U. C. M. Al- al diversity and the biosphere. Science 276: shington, DC: ASM Press.
smark, R. M. Podowski, A. K. Naslund, A.- 734-740. [R] Sive, H. L., R. M. Grainger and R. M. Harland
S. Erksson, H. H. Winkler and C. G. Kur- Shixing, Z. and C. Huineng. 1995. Mega-sco- (eds.). 1999. Early Development of Xenopus
land. 1998. The genome sequence of Ricket- pic multicellular organisms from the 1700- laevis: A Course Manual. Plainview, NY: Cold
tsia prowazekii and the origin of mitochon- million-year-old Tuanshanzi formation in the Spring Harbor Laboratory Press.
dria. Nature 396: 133-140. [P] Jixian area, north China. Science 270: 620- The Arabidopsis Genome Initiative. 2000. Analy-
Brocks, J. J., G. A. Logan, R. Buic k and R. E. 622. [P] sis of the genome sequence of the flowering
Summons. 1999. Archean molecular fossils Woese C. R., O. Kandler and M. L. Wheelis. plant Arabidopsis thaliana. Nature 408: 796–
and the early rise of eukaryotes. Science 285: 1990. Towards a natural system of organis- 815. [P]
1033-1036. [P] ms: Proposal for the domains Archae, Bacte- The C. elegans Sequencing Consortium. 1998.
Bult, C. J. and 39 others. 1996. Complete ge- ria, and Eucarya. Proc. Natl. Acad. Sci. USA Genome sequence of the nematode C. ele-
nome sequence of the methanogenic Archa- 87: 4576-4579. [P] gans: A platform for investigating biology.
eon, Methanococcus jannaschii. Science 273: Science 282: 2012-2018 [P]
1058-1073. [P] Thisse, C. and L. I. Zon. 2002. Organogenesis—
Cech, T. R. 1986. A model for the RNA-cat- Células como Modelos Experimentais heart and blood formation from the zebrafish
alyzed replication of RNA. Proc. Natl. Acad. point of view. Science 295:457-462. [R]
Adams, M.D., and 194 others. 2000. The ge-
Sci. USA 83:4360 4363. [P] Venter, J. C. and 273 others. 2001. The sequence
nome sequence of Drosophila melanogaster.
Crick, F. H. C. 1968. The origin of the genetic of the human genome. Science 291:1304-
Science 287: 2185–2195. [P].
code. J. Mol. Biol. 38: 367-379. [P] 1351. [P]
Blattner, F.R., G. Plunkett III., C. A. Bloch, N.
Darnell, J. E. and W. E Doolittle.1986. Specu-
T. Perna, V. Burland, M. Riley, J. Collado-
lations on the early course of evolution. Proc.
Vides, J. D. Glasner, C. K. Rode, G. F. Ferramentas da Biologia Celular
Natl. Acad. Sci. USA 83: 1271-1275. [P]
Mayhew, J. Gregor, N. W. Davis, H. A. Ki-
Doolittle, W. F. 1999. Phylogenetic classificati-
rkpatrick, M. A. Goeden, D. J. Rose, B. Mau Bowers, W. E. 1998. Christian de Duve and the
on and the universal tree. Science 284: 2124-
and Y. Shao. 1997. The complete genome discovery of lysosomes and peroxisomes.
2128. [R]
sequence of Escherichia coli K-12. Science Trends Cell Biol. 8: 330-333 [R]
Gesteland, R. F., T. R. Cech e J. F., Atkins (eds.).
277: 1453 1462. [P] Cairns, J., G. S. Stent and J. D. Watson (eds.).
1999. The RNA World. 2nd ed. Plainview,
Botstein, D., S. A. Chervitz and J. M. Cherry. 1992. Phage and the Origins of Molecular Bi-
NY: Cold Spring Harbor Laboratory Press.
1997. Yeast as a model organism. Science 277: ology. Plainview, NY: Cold Spring Harbor
Gilbert, W. 1986. The RNA world. Nature
1259-1260. [R] Laboratory Press.
319:618. [R]
Goffeau, A. and 15 others. 1996. Life with 6000 Chalfie, M. 1995. Green fluorescent protein.
Johnston W.K., P.J. Unrau, M.S. Lawrence,
genes. Science 274: 546-567. [P] Photochem. Photobiol. 62: 651–656. [R]
M.E. Glasner and D.P. Bartel. 2001. RNA-
Hamilton, B. A. and W. N. Frankel. 2001. Of Claude, A. 1975. The coming of age of the cell.
catalysed RNA polymerization: accurate and
mice and genome sequence. Cell 107:13– Science 189: 433-435. [R]
general RNA-templated primer extension.
16.[R] De Duve, C.1975. Exploring cells with a cen-
Science 292: 1319–1325. [P]
Hogan, B., R. Beddington, F. Costantini and trifuge. Science 189: 186-194. [R]
Joyce, G. F. 1989. RNA evolution and the ori-
E. Lacey. 1994. Manipulating the Mouse Em- Eagle, H. 1955. Nutrition needs of mammalian
gins of life. Nature 338: 217-224. [R]
bryo. 2nd. ed. Plainview, NY: Cold Spring cells in tissue culture. Science 235: 442-447.
Knoll, A. H. 1992. The early evolution of euka-
Harbor Laboratory Press. [P]
ryotes: A geological perspective. Science 256:
International Human Genome Sequencing Flint, S. J., L. W. Enquist, R. M. Krug, V. R.
622-627. [R]
Consortium. 2001. Initial sequencing and Racaniello and A. M. Skalka. 1999. Princi-
Margulis, L.1992. Symbiosis in Cell Evolution.
analysis of the human genome. Nature ples of Virology: Molecular Biology, Pathoge-
2nd. ed. New York: W. H. Freeman.
409:860–921.[P] nesis, and Control. Washington, DC: ASM
Miller, S. L. 1953. A production of amino acids
Maliga, P., D. F. Klessig, A. R. Cashmore, W. Press.
under possible primitive earth conditions.
Gruissem and J. E. Varner (eds.). 1995. Me- Graham, J. and D. Rickwood (ed.). 1997. Sub-
Science 117: 528-529. [P]
thods in Plant Molecular Biology. Plainview, cellular Fractionation: A Practical Approach.
Mojzsis, S. J., G. Arrhenius, K. D. McKee-
NY: Cold Spring Harbor Laboratory Press. New York: Oxford University Press.
gan, T. M. Harrison, A. P. Nutman and C.
Meyerowitz, E. M. 2002. Plants compared to Kam, Z., E. Zamir and B. Geiger. 2001. Pro-
R. L. Friend. 1996. Evidence for life on
development. Science 295: 1482–1485. [R] bing molecular processes in live cells by quan-](https://image.slidesharecdn.com/introduoabilogiacelular-130224145236-phpapp01/85/Introducao-a-biologia-celular-Leitura-complementar-38-320.jpg)
![40 / GEOFFREY M. COOPER • ROBERT E. HAUSMAN
titative multidimensional microscopy. Piston, D. W. 1999. Imaging living cells and Salmon, E. D. 1995. VE-DIC light microscopy
Trends Cell Biol. 11: 329–334. [R] tissues by two-photon excitation microsco- and the discovery of kinesin. Trends Cell Biol.
Lacey, A. J. (ed.) 1999. Light Microscopy in py. Trends Cell Biol. 9: 66-69 [R] 5:154 158. [R]
Biology: A Practical Approach. New York: Porter, K. R., A. Claude and E. F. Fullam. 1945. Spector, D. L., R. Goldman and L. Leinwand.
Oxford University Press. A study of tissue culture cells by electron mi- 1998. Cells: A Laboratory Manual. Plainview,
Palade, G. 1975. Intracellular aspects of the croscopy. J. Exp. Med. 81: 233 246. [P] NY: Cold Spring Harbor Laboratory Press.
process of protein synthesis. Science 189: Rous, P. 1911. A sarcoma of the fowl transmis- Tsien, R. Y. and A Miyawaki. 1998. Seeing the
347-358. [R] sible by an agent separable from the tumor machinery of live cells. Science 280: 1954-
cells. J. Exp. Med. 13:397411. [P] 1955. [R]](https://image.slidesharecdn.com/introduoabilogiacelular-130224145236-phpapp01/85/Introducao-a-biologia-celular-Leitura-complementar-39-320.jpg)

Este capítulo introduz a biologia celular e molecular. Discute a evolução das células a partir de um ancestral comum há bilhões de anos, incluindo a formação espontânea de moléculas orgânicas e o desenvolvimento do RNA auto-replicativo dentro de membranas de fosfolipídeos como a primeira célula. Também descreve as propriedades básicas e diferenças entre células procarióticas e eucarióticas.























































































