1) O relatório apresenta os resultados de um estudo sobre a pesca e aspectos reprodutivos do robalo-flecha na Ilha das Canárias no Delta do Parnaíba, Brasil.
2) Foram realizados monitoramento da pesca, coleta de amostras biológicas e análises histológicas para avaliar a sazonalidade da pesca, esforço de pesca, período reprodutivo e comprimento de primeira maturidade.
3) Os dados obtidos fornecem informações cruciais para o manejo sustentável
![RELATÓRIO FINAL – MONITORAMENTO DA PESCA, ESTUDO DOS ASPECTOS
REPRODUTIVOS E BIOMASSA DO ROBALO-FLECHA Centropomus undecimalis
(Bloch 1972) NA ILHA DAS CANÁRIAS
PROGRAMA PESCA PARA SEMPRE BRASIL_RESEX DELTA DO PARNAÍBA
UNIVERSIDADE FEDERAL DO PIAUÍ- UFPI
Dr. Cezar A. F. Fernandes
[Lab. de Bioecologia Pesqueira]
Dra. Francisca Edna de A. Cunha
[Lab. de Ictiologia]
10/07/2017](https://image.slidesharecdn.com/relatriofinalcezarfernandesednacunhasetembro2017-170930205851/85/Relatorio-Final-Robalo-2017-1-320.jpg)
![RELATÓRIO FINAL – MONITORAMENTO DA PESCA, ESTUDO DOS ASPECTOS
REPRODUTIVOS E BIOMASSA DO ROBALO-FLECHA Centropomus undecimalis
(Bloch 1972) NA ILHA DAS CANÁRIAS
PROGRAMA PESCA PARA SEMPRE BRASIL_RESEX DELTA DO PARNAÍBA
UNIVERSIDADE FEDERAL DO PIAUÍ- UFPI
Dr. Cezar A. F. Fernandes
[Lab. de Bioecologia Pesqueira]
Dra. Francisca Edna de A. Cunha
[Lab. de Ictiologia]
10/07/2017](https://image.slidesharecdn.com/relatriofinalcezarfernandesednacunhasetembro2017-170930205851/75/Relatorio-Final-Robalo-2017-1-2048.jpg)












![1/[1+exp (a+b*100*CP) ], onde MF é a fração dos indivíduos que se
encontram aptos a se reproduzirem (Beverton & Holt, 1956;
Shinozaki-Mendes et al., 2007). O ajuste dos pontos será realizado
através do método de máxima verossimilhança (maximun likelihood).
Para classificar o tipo de desenvolvimento ovocitário e o
padrão de regulação de fecundidade foram obtidas as frequências de
diâmetros dos ovócitos para cada estágio de desenvolvimento dos
ovócitos e para cada fase de maturidade sexual (Kjesbu et al.,
2003).
Para fecundidade foi adotada a metodologia adaptada de Hunter
(1985), Murua et al. (2003), Kjesbu et al. (2003) e Evans et al.
(2007), fecundidade por lote “Batch fecundity” através do método
gravimétrico, onde se retira 3 amostras de 0,01 g para cada fêmea
ativamente desovando, se conta os ovócitos hidratados e se usa a
seguinte fórmula abaixo:
F= ∑
Onde:
F= produto do peso da gônada e densidade dos ovócitos
Oi= número de ovócitos de cada porção
Wi= peso de cada porção do ovário
n= número de repetições de cada porção
Wovário= peso de cada ovário
6.5. Aplicação da análise estatística para dados da pesca e de
reprodução](https://image.slidesharecdn.com/relatriofinalcezarfernandesednacunhasetembro2017-170930205851/85/Relatorio-Final-Robalo-2017-14-320.jpg)





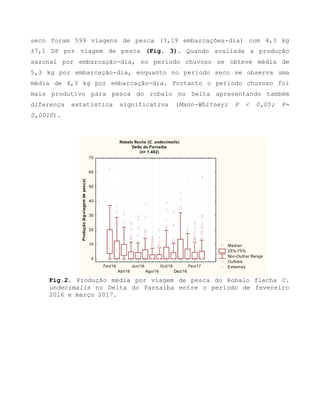
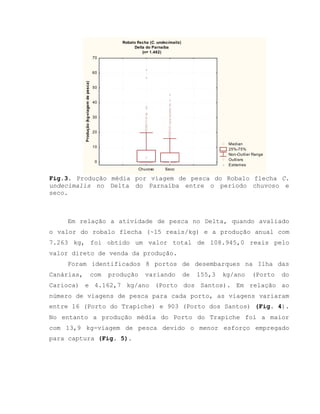
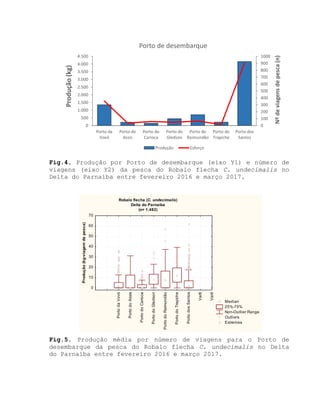
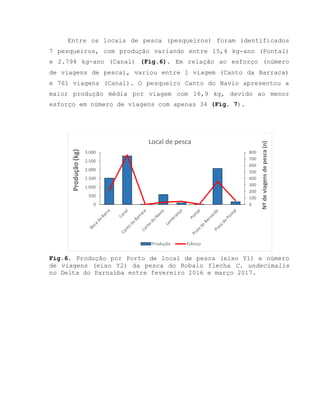
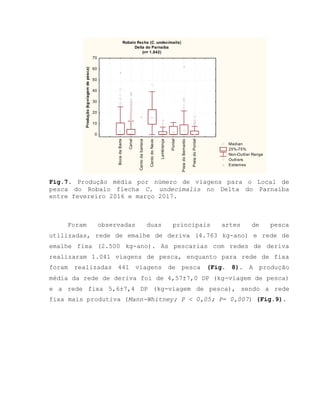
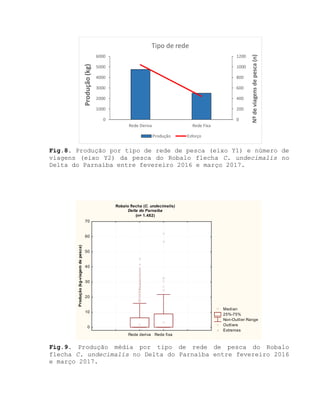
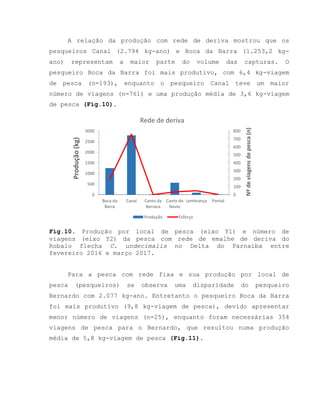
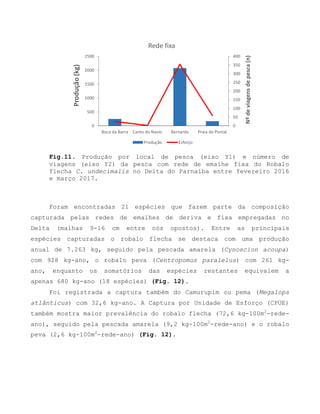
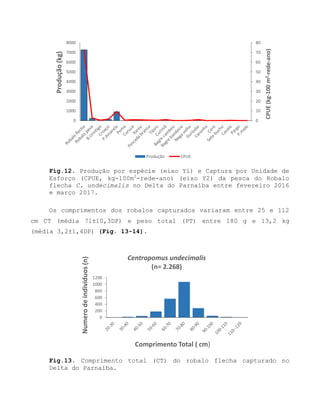
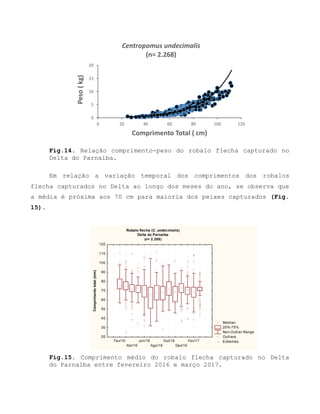
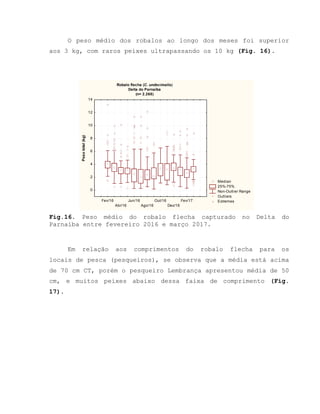
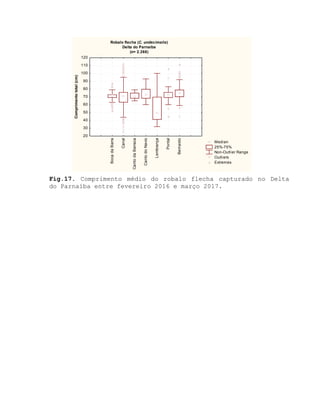


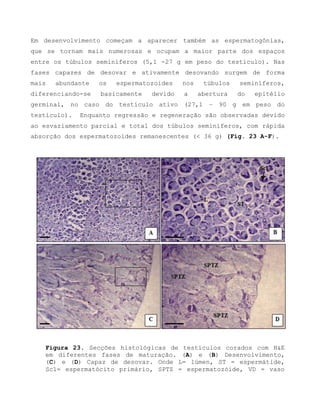
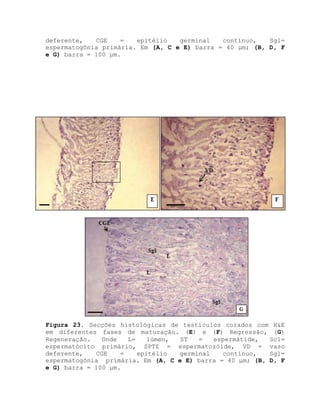


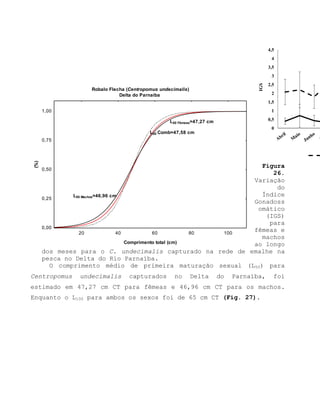







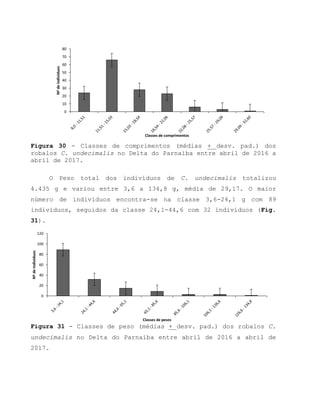





![Barroso, M.V; SOUZA, G. A; THOMÉ, J. C. A; LEITE JÚNIOR,
N.O; MOREIRA, L. M.P; SANGALIA, C; SALES, E. F; DURÃO, J.N.
2007. Estratégias de conservação das populações de robalos
Centropomus spp. na foz do Rio Doce, Linhares, Espírito
Santo, Brasil. Rev. Bras. de Agroecologia, Vol.2, No.2.
Begossi, A., Salivonchyk S.V., Araujo, L.G., Andreoli T.B.,
Clauzet, M. Martinelli C.M., Ferreira A.G.L., Oliveira
L.E.C., Silvano R.A.M. 2011. Ethnobiology of snappers
(Lutjanidae): target species and suggestions for management.
Journal of Ethnobiology and Ethnomedicine, 7:11.
Bittencourt, V. F. N. 2009. Proteção a Pesca do Robalo
(Centropomus parallelus) na Foz do Rio Mambucaba, Angra do
Reis-RJ. REVISTA - Educação Ambiental BE-597 / Volume 2,
38-43.
Brown-Peterson, N. J., D. M. Wyanski, F. Saborido-Rey, B. J.
Macewicz, & S. K. Lowerre-Barbieri. 2011. A standardized
terminology for describing reproductive development in
fishes. Marine and Coastal Fisheries: Dynamics, Management,
and Ecosystem Science [online serial], 3:52–70.
CAVALCANTE, C. C.: O uso de habitats costeiros como berçário
de peixes no Ceará. 2014. 71f.: Dissertação (mestrado) –
Universidade Federal do Ceará, Instituto de Ciências do Mar,
Programa de Pós-Graduação em Ciências Marinhas Tropicais.
Fortaleza, 2014.
Cerqueira, V. R. Cultivo de robalo: aspectos de reprodução,
larvicultura e engorda. Editora Autor, 94 p., Florianópolis,
2002.
Cullis-suzuki, Sarika; Mcallister, M.; Baker, P.;
Carruthers, T.; Tate, T.J. 2012. Red snapper discards in the
Gulf of Mexico: Fishermen’s perceptions following the
implementation of Individual Fishing Quotas. Marine Policy
36: 583–591.
DE MENDONÇA, Maisa Clari Farias Barbalho. Autoecologia do
camorim, Centropomus undecimalis (Bloch, 1792),(Perciformes:
Centropomidae) em ambiente hipersalino em Galinhos, RN,
Brasil. 2010.
Fernandes, C. A. F., Oliveira, P. G. V. de, Travassos, P. E.
P. F. & F. H. V. Hazin. 2012. Reproduction of the Brazilian
snapper, Lutjanus alexandrei (Moura & Lindeman, 2007)
(Perciformes: Lutjanidae), off the northern coast of](https://image.slidesharecdn.com/relatriofinalcezarfernandesednacunhasetembro2017-170930205851/85/Relatorio-Final-Robalo-2017-64-320.jpg)
![Pernambuco, Brazil. Neotropical Ichthyology, 10 (3): 587-
592.
Frédou, T., Ferreira, B.P., Letourneur, Y., 2009. Assessing
the stocks of the primary snappers caught in Northeastern
Brazilian reefs systems. 1: Traditional modeling approaches.
Fisheries Research. 99 (2009) 90-96.
Ganias, K. 2009. Linking sardine spawning dynamics to
environmental variability. Estuarine, Coastal, and Shelf
Science 84:402–408.
Guzzi, A . Biodiversidade do Delta do Parnaíba: litoral
piauiense. / Anderson Guzzi. – org. Parnaíba: EDUFPI, 2012.
466p.
Hardardottir, K., Kjesbu, O.S., & G. Marteinsdottir. 2001.
Relationship between atresia, fish size and condition in
Icelandic cod (Gadus morhua L.). ICES CM2001/J:19.
HUBER, C. G. Habitat preferences of juvenile Common Snook in
the lower Rio Grande, Texas. 2007. Tese de Doutorado.
Master’s thesis. Texas Tech University, Lubbock.
Hunter. J. R. & B. Macewicz. 1985c. Measurement of spawning
frequency in multiple spawning fishes. In: Lasker, R. (Ed).
An egg production method for estimating spawning biomass of
pelagic fish: application to the Northern Anchovy, Engraulis
mordax. NOAA Technical Report NMFS, 36: 79-93.
Lowerre-Barbieri, S. K., K. Ganias, F. Saborido-Rey, H.
Murua, & J. R. Hunter. 2011b. Reproductive timing in marine
fishes: variability, temporal scales, and methods. Marine
and Coastal Fisheries: Dynamics, Management, and Ecosystem
Science [online serial], 3:71–91.
Machado, M. R. F. 2011. Caracterização Morfológica e
Bioquímica do Sistema Digestório e Identificação por
Isótopos Estáveis de Robalo Peva e Flexa Selvagens e de
Cativeiro. Tese apresentada ao Programa de Pós - graduação
em Aquicultura da UNESP – Campus de Jaboticabal, como parte
das exigências para obtenção do título de Doutor(a) em
Aquicultura. Jaboticabal São Paulo – Brasil 2011.](https://image.slidesharecdn.com/relatriofinalcezarfernandesednacunhasetembro2017-170930205851/85/Relatorio-Final-Robalo-2017-65-320.jpg)















































