Baixado 256 vezes









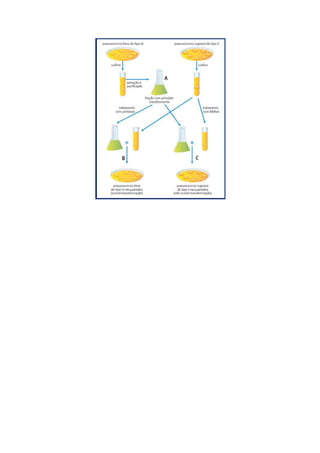

O documento descreve a estrutura do DNA em três níveis: primária, secundária e terciária. A estrutura primária consiste em nucleotídeos unidos por ligações fosfodiéster. A estrutura secundária forma uma dupla-hélice com dois filamentos complementares mantidos juntos por pontes de hidrogênio entre bases. A estrutura terciária inclui diferentes configurações tridimensionais da dupla-hélice.











































































