1) O documento discute a evolução dos grandes grupos de animais e como os mecanismos genéticos do desenvolvimento influenciaram essa evolução.
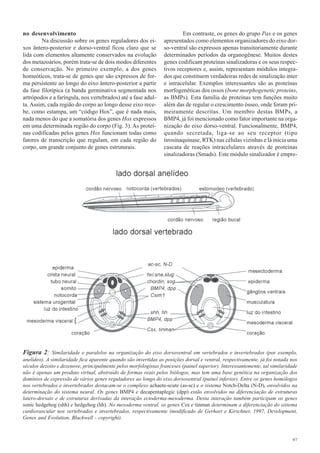
2) Muitos genes reguladores do desenvolvimento são altamente conservados entre os metazoários e definiram as identidades dos segmentos corporais durante a evolução dos filos.
3) Pequenas mudanças nesses genes reguladores poderiam ter gerado a diversidade nos planos corporais observados nos 35 filos animais atuais.