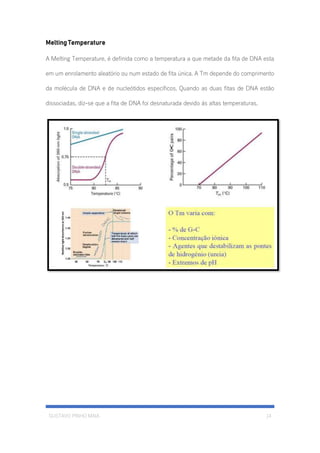
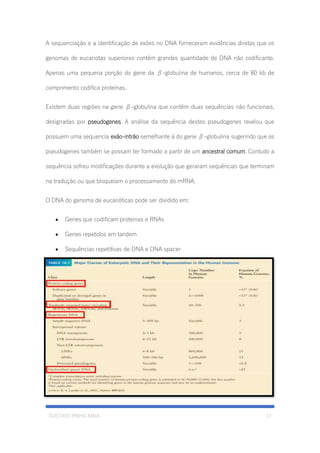
O documento descreve a estrutura e constituição da célula eucariótica, incluindo o núcleo, cromatina, cromossomos e DNA. Detalha a estrutura do invólucro nuclear, poros nucleares, cromatina, cromossomos e o modelo de Watson e Crick para a estrutura em dupla hélice do DNA.