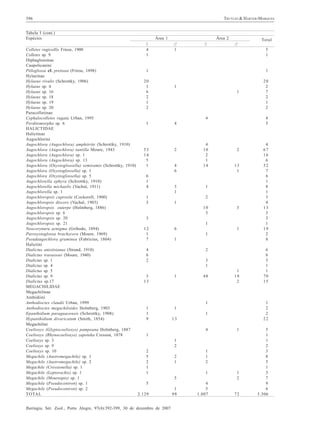
O estudo avaliou a comunidade de abelhas em duas áreas florestais do Parque Estadual de Itapuã no Rio Grande do Sul. Foram amostradas 3,306 abelhas de 95 espécies ao longo de um ano. A família Apidae foi a mais abundante e diversa, seguida por Halictidae. A atividade das abelhas reduziu em março e abril, quando a precipitação foi menor e as temperaturas maiores.