Baixado 28 vezes



![Programa de Pós-Graduação em Ecologia – CCB - UFSC
Copyright © 2010 – PPG Ecologia UFSC
Impresso no Brasil / Printed in Brazil
Diagramação: Mauricio Cantor, Luis Carlos Pinto de Macedo-Soares,
Natalia Hanazaki
Foto da capa: Gabriela Corso da Silva, Heraclides hectorides.
Revisão: Autores e organizadores
Catalogação na fonte elaborada na DECTI da Biblioteca da UFSC
[ficha catalográfica]
Programa de Pós-Graduação em Ecologia
Centro de Ciências Biológicas UFSC
Campus Universitário Trindade
Florianópolis, SC, Brasil
CEP 88010-970 poseco@ccb.ufsc.br
www.poseco.ufsc.br
E19 Ecologia de campo na Lagoa do Peri 2009 / organizadores
Mauricio Cantor, Luis Carlos Pinto Macedo-Soares,
Natalia Hanazaki. – 1. ed. - Florianópolis : Universidade
Federal de Santa Catarina, 2010.
193 p. : il., grafs., tabs., mapas
Inclui referências.
ISBN: 9788574261058
1. Parque Municipal da Lagoa do Peri (SC) – Ecologia.
2. Ecologia lacustre. I. Cantor, Maurício. II. Macedo-
Soares, Luís Carlos Pinto de. III. Hanazaki, Natália.
CDU: 577.4](https://image.slidesharecdn.com/ecologiadecampov22009-140805120745-phpapp01/85/Ecologia-de-Campo-2009-POSECO-UFSC-3-320.jpg)





















































































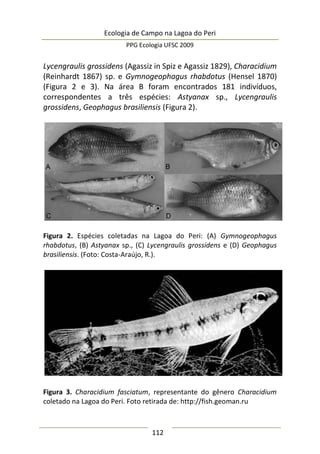

















































![Ecologia de Campo na Lagoa do Peri
PPG Ecologia UFSC 2009
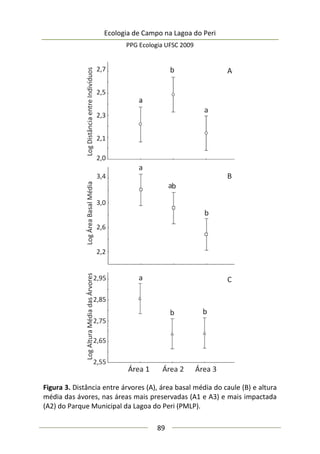
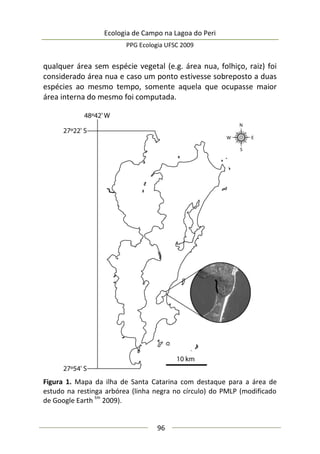
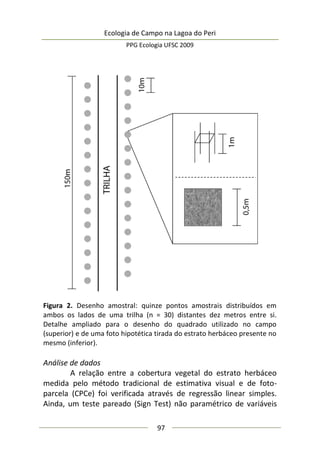
173
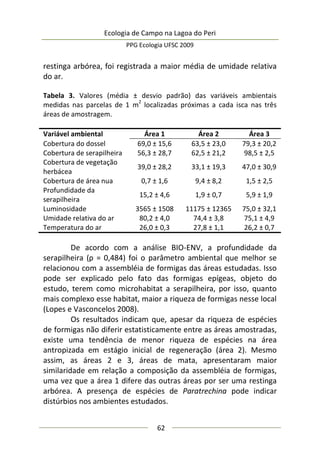
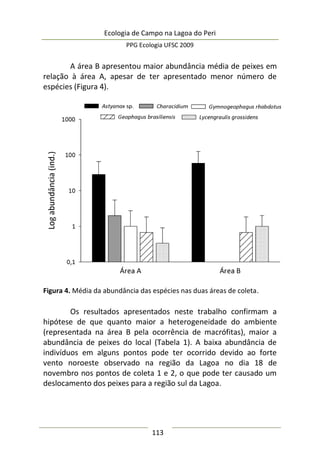
Análise de dados
Uma matriz de correlação de Pearson foi construída com as
variáveis de complexidade de habitat e abundância de predadores
por umbela, enquanto a significância entre os pares foi verificada
por teste-t unicaudal. Correlação de Spearman foi utilizada para
analisar a relação entre densidade visualmente estimada da
vegetação e abundância de predadores. Uma regressão linear
simples foi utilizada para descrever o comportamento da
abundância de aracnídeos em função da variação dos descritores de
complexidade estrutural. Análise de variância (ANOVA) e
distribuição de Fischer-Snedecor foram aplicadas para testar a
significância da regressão. O modelo foi ajustado mediante análise
de resíduos, excluindo outliers fora do intervalo [-2;+2]. A
normalidade da distribuição de todos os dados foi previamente
testada pelo teste de Shapiro-Wilk. Para todas as análises foi
considerado o nível de significância de α = 0,05 (Zar 2007).
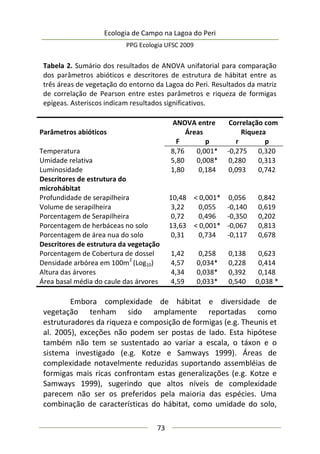
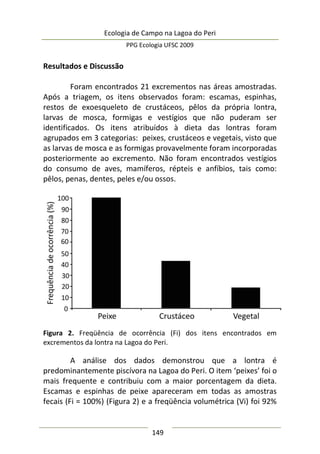
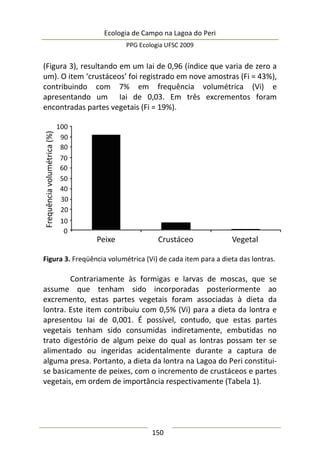
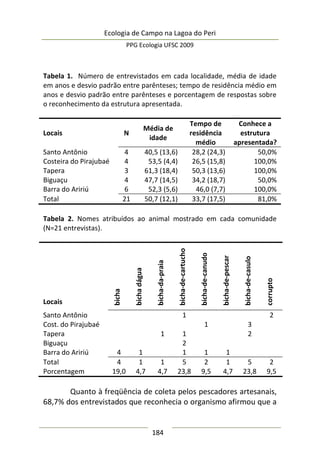
Resultados e Discussão
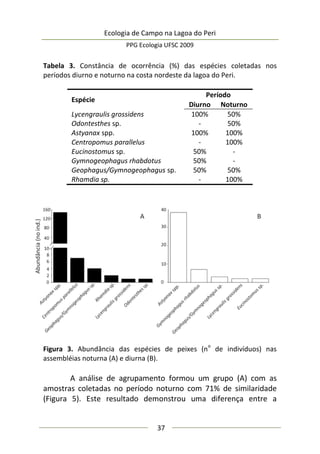
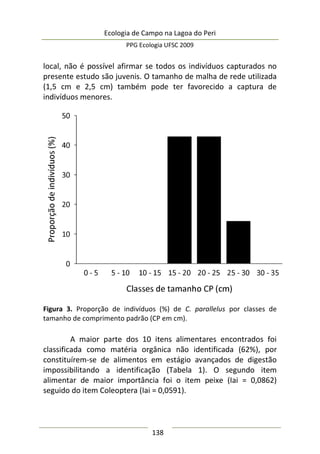
Neste estudo foram encontrados 158 aracnídeos não
formadores de teia, a maioria pertencente à Família Thomisidae,
morfoidentificados pelos dois primeiros pares de pernas maiores,
mais robustos e dispostos lateralmente. Estes aracnídeos alojaram-
se em 312 inflorescências em forma de umbela de Actinocephalus
polyanthus ( = 1,33 indivíduos/umbela ± 0,50 DP).
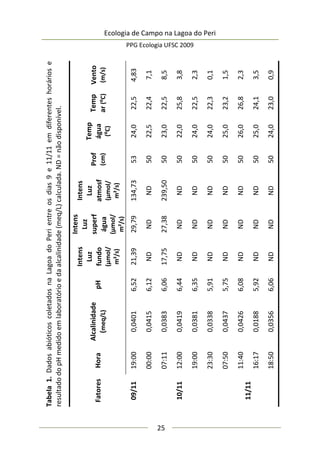
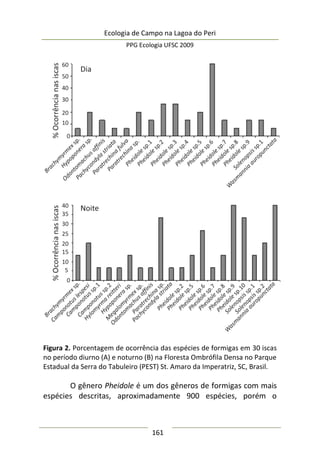
Complexidade de habitat é um fator importante na
determinação da diversidade, distribuição de espécies e abundância
de aranhas (e.g. Hatley e MacMahon 1980, Halaj et al. 1998). Uma
série de características de um tipo de habitat utilizado por aranhas
de tocaia foi avaliada neste estudo, porém somente a altura da
estrutura reprodutiva total da planta, o diâmetro e volume médio
de suas umbelas apresentaram correlação significativa com a
abundância de aracnídeos (Tabela 1). A presença de um organismo](https://image.slidesharecdn.com/ecologiadecampov22009-140805120745-phpapp01/85/Ecologia-de-Campo-2009-POSECO-UFSC-174-320.jpg)


![Ecologia de Campo na Lagoa do Peri
PPG Ecologia UFSC 2009
176
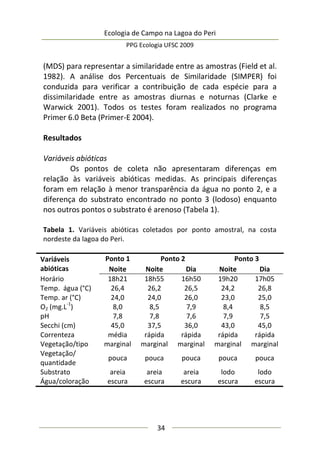
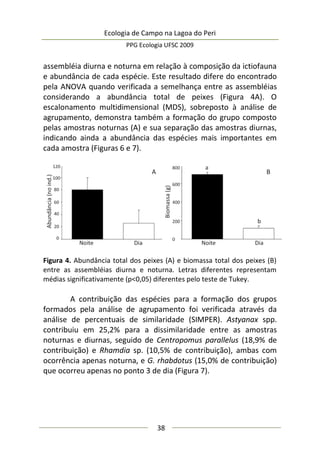
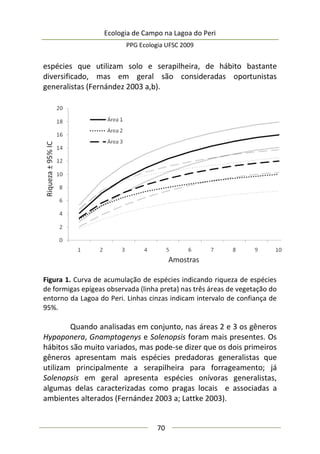
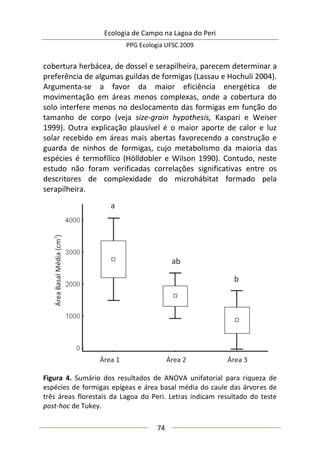
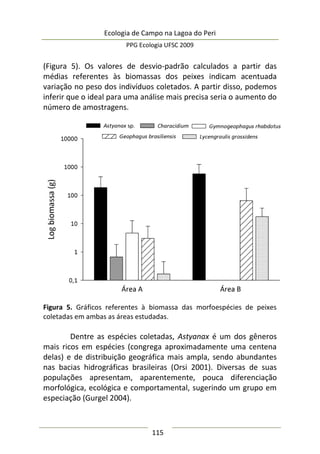
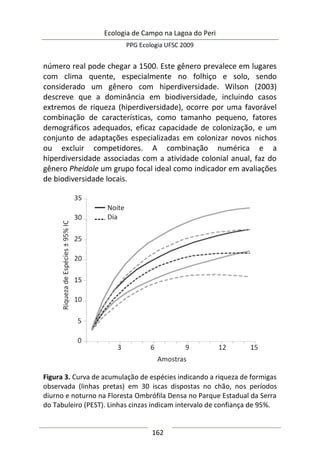
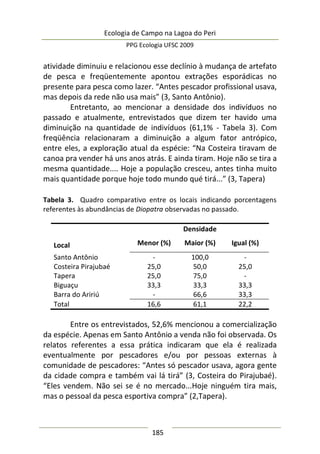
A moderada correlação entre abundância de predadores e
diâmetro da umbela permitiu estimar o comportamento da
abundância de predadores: a relação entre elas foi positiva e
significativa (F(1,25) = 9,47; p = 0,005; EPestimativa = 3,43), representada
no modelo da Figura 3, melhor ajustado com a exclusão de três
outliers. Este modelo sugere que maiores umbelas suportam mais
aranhas de tocaia. Embora o tamanho amostral aqui empregado
seja razoável e adequado para regressão linear simples, o potencial
de interpretação desta análise é moderado, pois existe um
significativo erro embutido que torna o modelo impreciso. A
maioria dos valores de diâmetro de umbela se concentra no
intervalo [6,5-8,0 cm] (Figura 3), sugerindo que a amostragem pode
não ter sido suficiente para detectar equitativamente toda
amplitude de variação.
Figura 3. Regressão linear entre diâmetro médio da inflorescência
(umbela) de A. polyanthus e abundância de aracnídeos predadores por
umbela.](https://image.slidesharecdn.com/ecologiadecampov22009-140805120745-phpapp01/85/Ecologia-de-Campo-2009-POSECO-UFSC-177-320.jpg)

















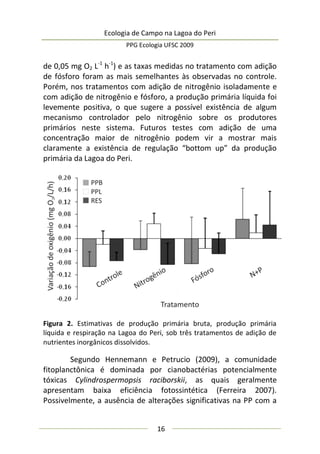
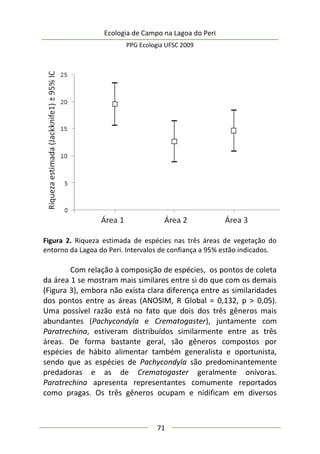
Este estudo avaliou a produção primária natural em uma área marginal da Lagoa do Peri e como o incremento de nutrientes (nitrogênio e fósforo) poderia influenciar esta produção. Amostras de água foram coletadas em garrafas de incubação para estimar a taxa de produção primária bruta por fotossíntese. Os resultados indicaram que o fósforo é um nutriente limitante na Lagoa do Peri e que o incremento deste elemento aumentou a produção primária no local.

























































![Plano De Disciplina Ecologia [1]](https://cdn.slidesharecdn.com/ss_thumbnails/planodedisciplinaecologia1-091202071746-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)


























