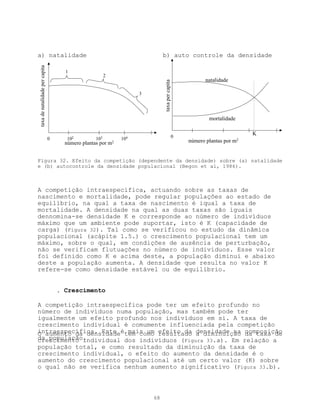
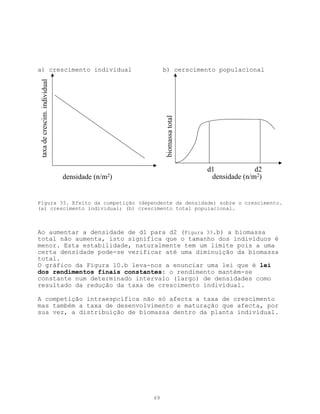
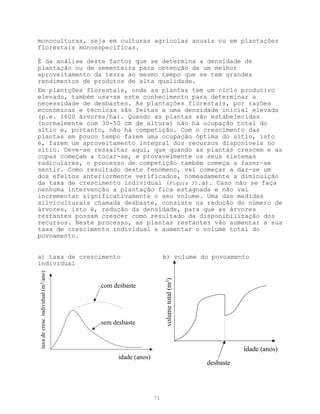
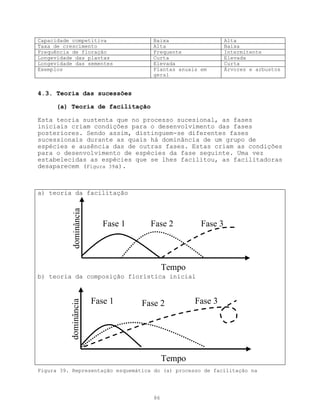
Este documento apresenta uma introdução sobre ecologia, definindo o termo e descrevendo os principais níveis de estudo. Aborda também a vegetação, definindo o termo e descrevendo a natureza e evolução da vegetação ao longo da história geológica da Terra. Por fim, fornece uma definição de ecologia como ciência descritiva, explicativa e preditiva.










































































































![Aulas De Ecologia E Meio Ambiente1[1]](https://cdn.slidesharecdn.com/ss_thumbnails/aulasdeecologiaemeioambiente11-091202060450-phpapp01-thumbnail.jpg?width=640&height=640&fit=bounds)


























































