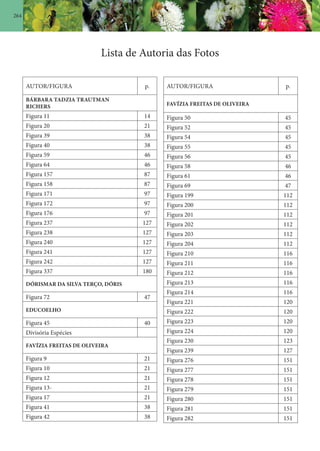
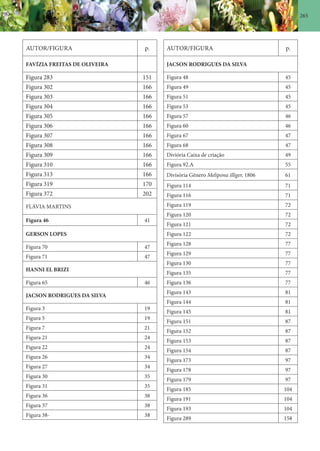
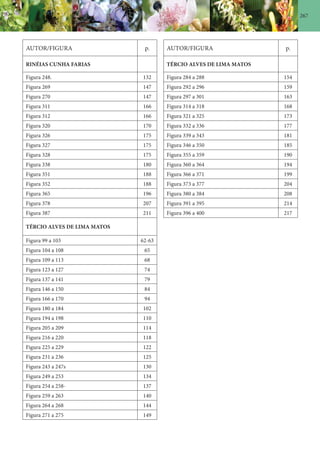
Baixado 138 vezes




















































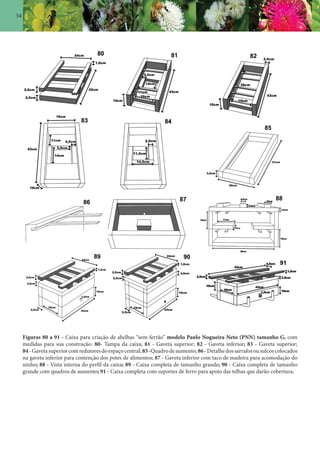

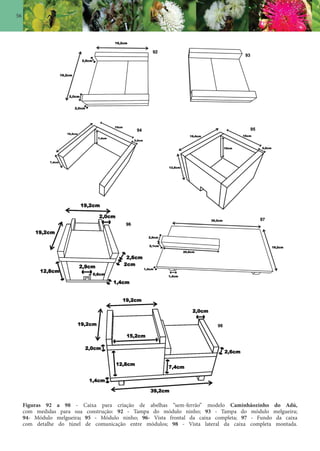







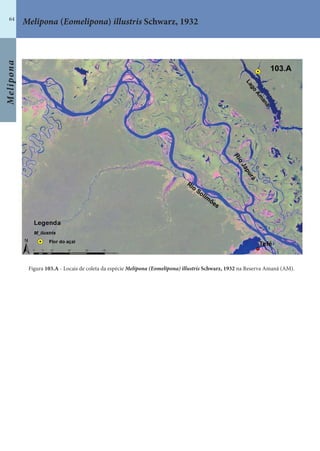








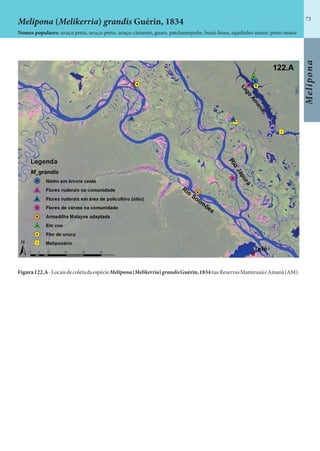




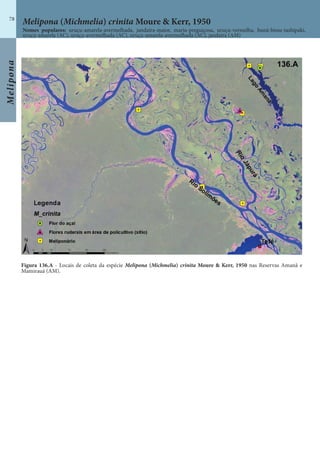






















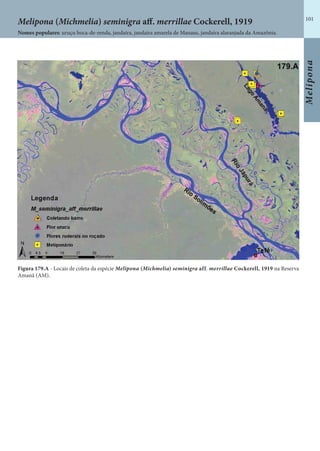





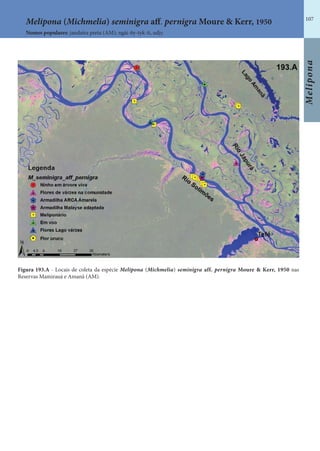




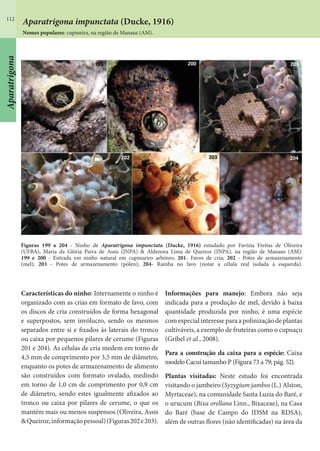













































































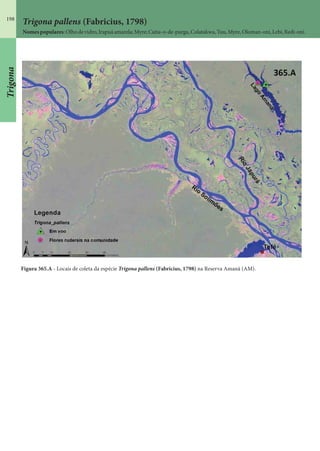














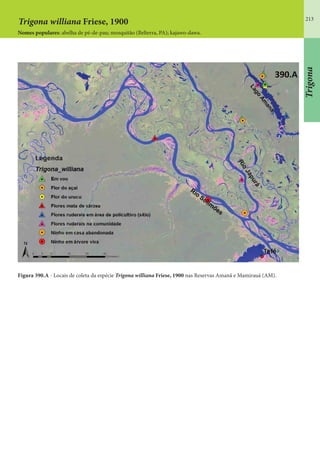
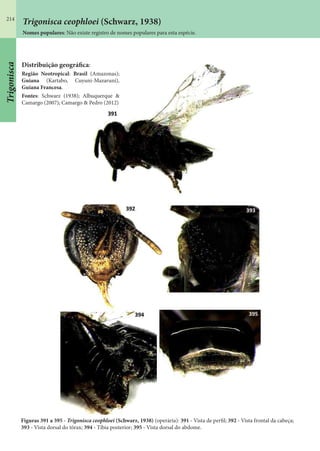


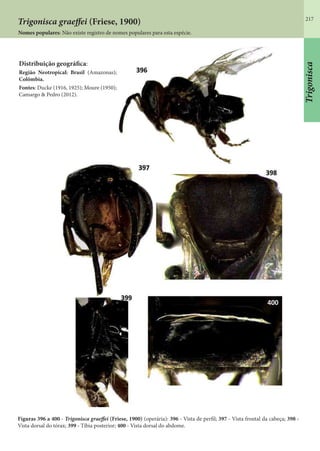


























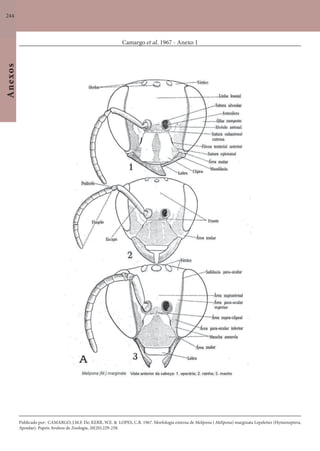
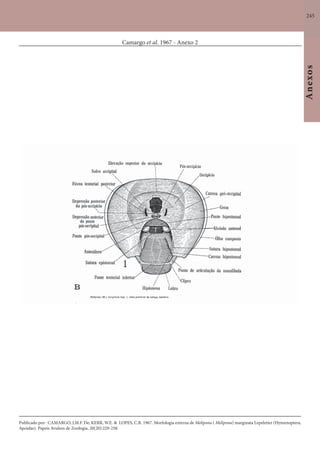
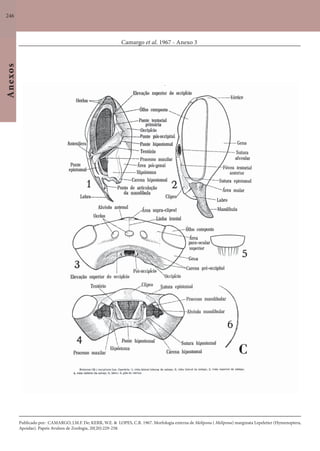
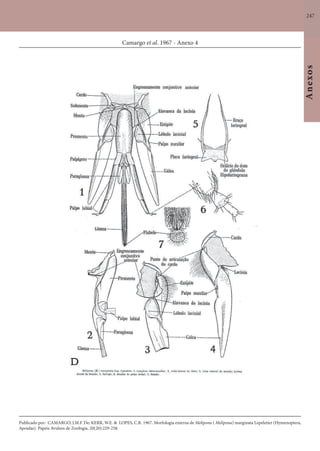
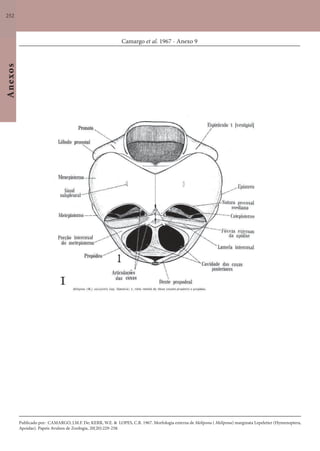
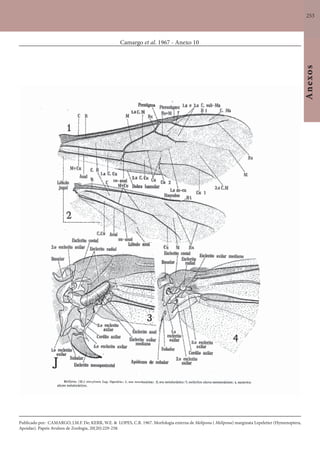
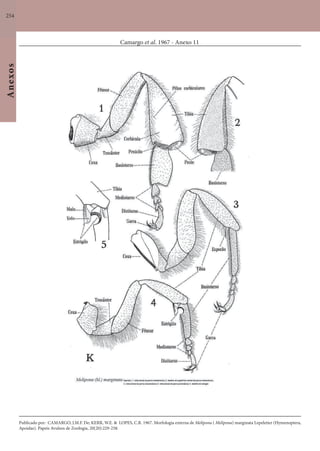
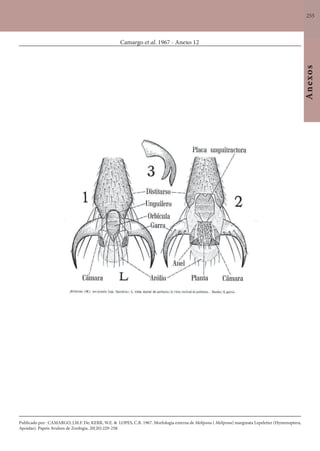
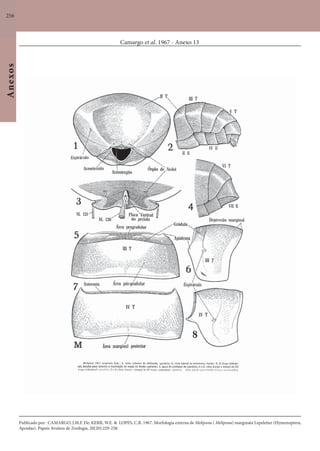
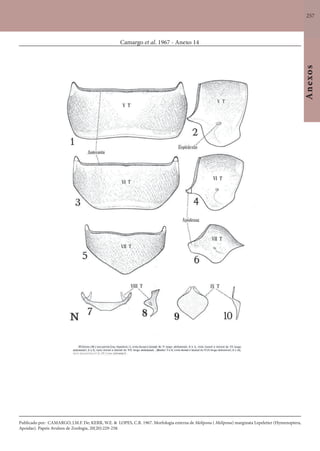
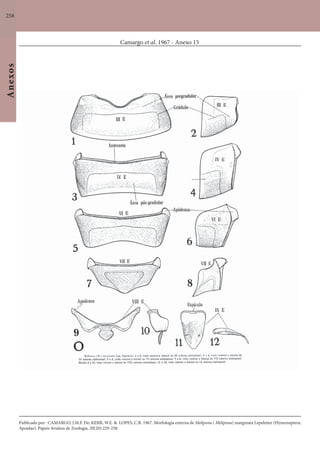
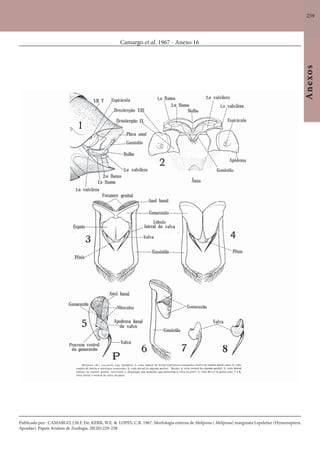







Este documento é um guia ilustrado das abelhas sem ferrão das Reservas Amanã e Mamirauá no Amazonas, Brasil. O guia descreve as espécies de abelhas encontradas nas reservas, incluindo informações sobre identificação, hábitos de nidificação e flores visitadas. Além disso, fornece instruções sobre a criação de caixas para o manejo dessas abelhas e uma chave de identificação para as espécies.








![(Mapa Mental) SUS - LEI 8080 - [Da Organização, da Direção e da Gestão]](https://cdn.slidesharecdn.com/ss_thumbnails/be25fa50d51c11e7914c118552ea79f4-171129154937-thumbnail.jpg?width=640&height=640&fit=bounds)










![SUS lei 8080 [Disposições Preliminares; Definição do Sistema Único de Saúde]](https://cdn.slidesharecdn.com/ss_thumbnails/mapamentallei8080-susparte1-171128213609-thumbnail.jpg?width=640&height=640&fit=bounds)































































