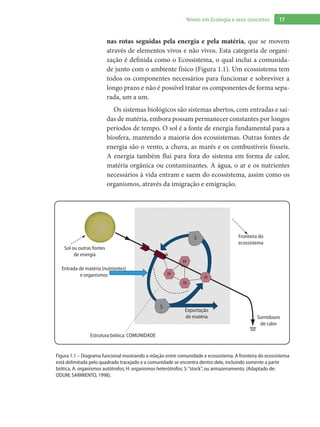
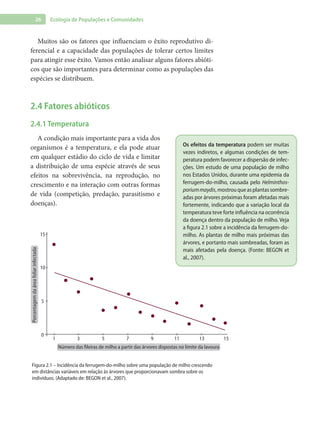
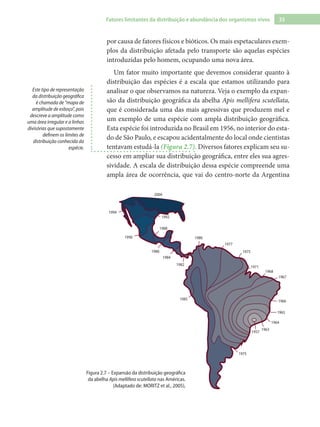
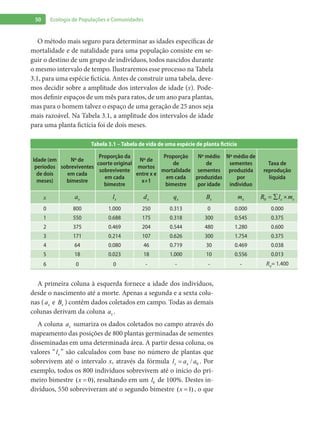
Baixado 1.880 vezes



























































![64 Ecologia de Populações e Comunidades
Considerando que 1t tN N+ − expressa uma mudança, podemos
representar essa mudança por N∆ [Este símbolo (∆ ) é a letra gre-
ga delta e lemos então “delta N”], que representa a mudança no
número de indivíduos. Esse símbolo ∆ poderia ser usado também
para representar uma mudança entre o tempo 0t = e 1t = , ou seja,
0t e 1t , t∆ .
Considerando essas duas taxas de mudanças, ou seja, da mu-
dança do número de indivíduos ( N∆ ) e da variação de tempo
( t∆ ), agora poderíamos calcular a taxa média de mudança no nú-
mero de organismos por tempo, ou seja, dividir as duas taxas de
mudança, /N t∆ ∆ . Essa taxa resultante, entretanto, é uma média,
e poderíamos nos perguntar qual seria uma taxa instantânea de
crescimento, ou seja, quando t∆ é tão pequeno que teoricamente
teríamos um valor de crescimento num momento qualquer da his-
tória de vida daquela população que estamos estudando.
Vamos considerar então que o crescimento da população que es-
tamos querendo modelar é contínuo e que queremos achar um va-
lor dentro de um momento muito pequeno de tempo: isso significa
que o intervalo de tempo entre tN e 1tN + é infinitamente pequeno.
Para fazer isso matematicamente, é preciso derivar a equação aci-
ma e substituir o símbolo ∆ pela letra “d”, o que equivale a dizer:
/dN dt B M= −
Lê-se estritamente: a derivada de N pelo tempo t é igual a
nascimentos menos mortes, porém este é um termo técnico e
pouco usual. Você deve entender que é a maneira de represen-
tar uma taxa de mudança, neste caso a mudança do número de
indivíduos pelo tempo. Veja neste caso como é importante ter
muito claro o que são, quais são, e quantos são os indivíduos
da população em estudo.
Observe que /dN dt é uma maneira de expressar que há uma
mudança no número de indivíduos por unidade de tempo.
Lembre-se que no capítulo anterior vimos a diferença entre nú-
mero de nascimentos e taxa de natalidade, e número de mortes e](https://image.slidesharecdn.com/livro-ecologia-de-populacoes-e-comunidades-130526143842-phpapp01/85/Livro-Ecologia-de-Populacoes-e-Comunidades-64-320.jpg)






















































O documento apresenta um capítulo sobre níveis em Ecologia e seus conceitos. Ele define Ecologia e seus principais níveis de organização, com ênfase na diferença entre Ecologia de Populações e Ecologia de Comunidades. Também define população como unidade de estudo em Ecologia de Populações, focando em aspectos como abundância, distribuição, sobrevivência e reprodução.























































































