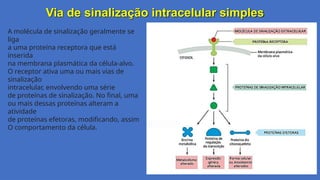
Via de sinalizaçãointracelular simples
Gláucia Ma2rques
A molécula de sinalização geralmente se
liga
a uma proteína receptora que está
inserida
na membrana plasmática da célula-alvo.
O receptor ativa uma ou mais vias de
sinalização
intracelular, envolvendo uma série
de proteínas de sinalização. No final, uma
ou mais dessas proteínas alteram a
atividade
de proteínas efetoras, modificando, assim
O comportamento da célula.
3.
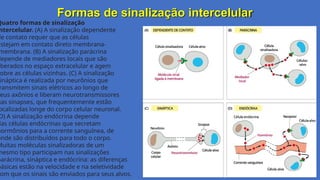
Formas de sinalizaçãointercelular
Gláucia Ma3rques
Quatro formas de sinalização
ntercelular. (A) A sinalização dependente
de contato requer que as células
estejam em contato direto membrana-
membrana. (B) A sinalização parácrina
depende de mediadores locais que são
iberados no espaço extracelular e agem
obre as células vizinhas. (C) A sinalização
ináptica é realizada por neurônios que
ransmitem sinais elétricos ao longo de
eus axônios e liberam neurotransmissores
nas sinapses, que frequentemente estão
ocalizadas longe do corpo celular neuronal.
D) A sinalização endócrina depende
das células endócrinas que secretam
hormônios para a corrente sanguínea, de
onde são distribuídos para todo o corpo.
Muitas moléculas sinalizadoras de um
mesmo tipo participam nas sinalizações
parácrina, sináptica e endócrina: as diferenças
básicas estão na velocidade e na seletividade
om que os sinais são enviados para seus alvos.
4.
Moléculas de sinalizaçãoextracelular se ligam aos
receptores de superfície e intracelulares.
Gláucia Ma4rques
Ligação de moléculas de sinalização extracelular
aos receptores de superfície e intracelular:
(A) A maioria das moléculas de sinalização é
hidrofílica e, por isso, incapaz de atravessar a
membrana da célula-alvo, elas se ligam a
receptores de superfície que, por sua vez,
geram sinais no interior da célula-alvo.
(B) Algumas moléculas de sinalização pequenas se
difundem através da membrana plasmática e se
ligam a proteínas receptoras no interior da
célula-alvo (citosol ou núcleo). Muitas dessas
moléculas pequenas são hidrofóbicas e pouco
solúveis em soluções aquosas, por isso, são
transportadas, na corrente sanguínea ou em
outros fluidos extracelulares, ligadas a proteínas
carreadoras, das quais se dissociam antes de
entrar na célula-alvo
5.
Moléculas de sinalizaçãoextracelular se ligam aos
receptores de superfície e intracelulares.
Gláucia Ma5rques
Receptores de Superfície Celular:
• Moléculas hidrofílicas (ex.: hormônios peptídicos,
neurotransmissores) não conseguem atravessar
a membrana plasmática.
• Elas se ligam a proteínas receptoras
específicas na superfície da célula-alvo
(membrana plasmática).
• Essa ligação desencadeia uma cascata de
sinalização intracelular (ex.: ativação de proteínas
G, enzimas ou segundos mensageiros).
Receptores Intracelulares
• Moléculas hidrofóbicas (ex.: hormônios
esteroides, tireoidianos) atravessam a membrana
plasmática livremente.
• No citosol, podem se ligar a proteínas
carreadoras (ex.: proteínas de ligação a
hormônios) que as auxiliam no transporte até o
núcleo.
• No núcleo, ligam-se a receptores intracelulares,
Diferentes respostas induzidaspelo
neurotransmissor acetilcolina.
(B) Célula Marca-Passo Cardíaca
• Receptor: Muscarínico (tipo M₂), acoplado à proteína G
inibitória (Gi).
• Mecanismo:
• A ligação da ACh ao receptor M₂ ativa a proteína Gi,
que inibe a adenilil ciclase, reduzindo os níveis de
AMP cíclico (cAMP).
• Isso abre canais de potássio (K⁺), causando
hiperpolarização da célula.
• Resultado: Redução na frequência de disparo do nó
sinoatrial, diminuindo a velocidade de contração
cardíaca (efeito parassimpático).
8.
Diferentes respostas induzidaspelo
neurotransmissor acetilcolina.
(C) Célula da Glândula Salivar
• Receptor: Muscarínico (tipo M₃), acoplado à proteína G
estimulatória (Gq).
• Mecanismo:
• A ligação da ACh ao receptor M₃ ativa a fosfolipase C
(PLC), que cliva PIP₂ em IP₃ e diacilglicerol (DAG).
• O IP₃ libera cálcio (Ca²⁺) do retículo endoplasmático,
enquanto o DAG ativa a proteína-cinase C (PKC).
• Resultado: Secreção de saliva devido à exocitose de
grânulos salivares.
9.
Diferentes respostas induzidaspelo
neurotransmissor acetilcolina.
(D) Célula Muscular Esquelética
• Receptor: Nicotínico (tipo Nm), um canal iônico ligante-
dependente.
• Mecanismo:
• A ligação da ACh ao receptor nicotínico abre o canal,
permitindo a entrada de Na⁺ e saída de K⁺.
• A despolarização resultante gera um potencial de ação na
membrana da célula muscular.
• Resultado: Contração muscular rápida via liberação de
Ca²⁺ do retículo sarcoplasmático.
10.
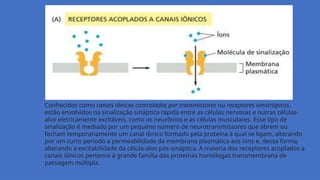
Conhecidos como canaisiônicos controlados por transmissores ou receptores ionotrópicos,
estão envolvidos na sinalização sináptica rápida entre as células nervosas e outras células-
alvo eletricamente excitáveis, como os neurônios e as células musculares. Esse tipo de
sinalização é mediado por um pequeno número de neurotransmissores que abrem ou
fecham temporariamente um canal iônico formado pela proteína à qual se ligam, alterando
por um curto período a permeabilidade da membrana plasmática aos íons e, dessa forma,
alterando a excitabilidade da célula-alvo pós-sináptica. A maioria dos receptores acoplados a
canais iônicos pertence à grande família das proteínas homólogas transmembrana de
passagem múltipla.
11.
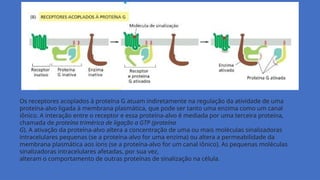
Os receptores acopladosà proteína G atuam indiretamente na regulação da atividade de uma
proteína-alvo ligada à membrana plasmática, que pode ser tanto uma enzima como um canal
iônico. A interação entre o receptor e essa proteína-alvo é mediada por uma terceira proteína,
chamada de proteína trimérica de ligação a GTP (proteína
G). A ativação da proteína-alvo altera a concentração de uma ou mais moléculas sinalizadoras
intracelulares pequenas (se a proteína-alvo for uma enzima) ou altera a permeabilidade da
membrana plasmática aos íons (se a proteína-alvo for um canal iônico). As pequenas moléculas
sinalizadoras intracelulares afetadas, por sua vez,
alteram o comportamento de outras proteínas de sinalização na célula.
12.
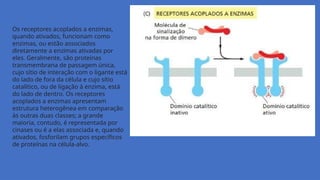
Os receptores acopladosa enzimas,
quando ativados, funcionam como
enzimas, ou estão associados
diretamente a enzimas ativadas por
eles. Geralmente, são proteínas
transmembrana de passagem única,
cujo sítio de interação com o ligante está
do lado de fora da célula e cujo sítio
catalítico, ou de ligação à enzima, está
do lado de dentro. Os receptores
acoplados a enzimas apresentam
estrutura heterogênea em comparação
às outras duas classes; a grande
maioria, contudo, é representada por
cinases ou é a elas associada e, quando
ativados, fosforilam grupos específicos
de proteínas na célula-alvo.
13.
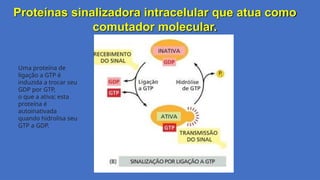
Proteínas sinalizadora intracelularque atua como
comutador molecular.
Uma proteína de
ligação a GTP é
induzida a trocar seu
GDP por GTP,
o que a ativa; esta
proteína é
autoinativada
quando hidrolisa seu
GTP a GDP.
14.
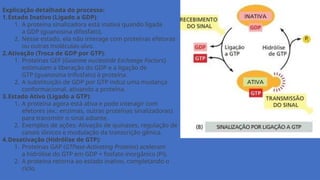
Explicação detalhada doprocesso:
1.Estado Inativo (Ligado a GDP):
1. A proteína sinalizadora está inativa quando ligada
a GDP (guanosina difosfato).
2. Nesse estado, ela não interage com proteínas efetoras
ou outras moléculas-alvo.
2.Ativação (Troca de GDP por GTP):
1. Proteínas GEF (Guanine nucleotide Exchange Factors)
estimulam a liberação do GDP e a ligação de
GTP (guanosina trifosfato) à proteína.
2. A substituição de GDP por GTP induz uma mudança
conformacional, ativando a proteína.
3.Estado Ativo (Ligado a GTP):
1. A proteína agora está ativa e pode interagir com
efetores (ex.: enzimas, outras proteínas sinalizadoras)
para transmitir o sinal adiante.
2. Exemplos de ações: Ativação de quinases, regulação de
canais iônicos e modulação da transcrição gênica.
4.Desativação (Hidrólise de GTP):
1. Proteínas GAP (GTPase-Activating Proteins) aceleram
a hidrólise do GTP em GDP + fosfato inorgânico (Pi).
2. A proteína retorna ao estado inativo, completando o
ciclo.
15.
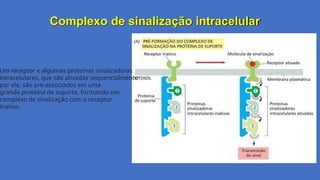
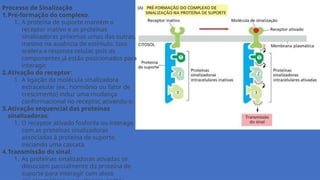
Complexo de sinalizaçãointracelular
Um receptor e algumas proteínas sinalizadoras
intracelulares, que são ativadas sequencialmente
por ele, são pré-associados em uma
grande proteína de suporte, formando um
complexo de sinalização com o receptor
inativo.
16.
Processo de Sinalização
1.Pré-formaçãodo complexo:
1. A proteína de suporte mantém o
receptor inativo e as proteínas
sinalizadoras próximas umas das outras,
mesmo na ausência de estímulo. Isso
acelera a resposta celular, pois os
componentes já estão posicionados para
interagir.
2.Ativação do receptor:
1. A ligação da molécula sinalizadora
extracelular (ex.: hormônio ou fator de
crescimento) induz uma mudança
conformacional no receptor, ativando-o.
3.Ativação sequencial das proteínas
sinalizadoras:
1. O receptor ativado fosforila ou interage
com as proteínas sinalizadoras
associadas à proteína de suporte,
iniciando uma cascata.
4.Transmissão do sinal:
1. As proteínas sinalizadoras ativadas se
dissociam parcialmente da proteína de
suporte para interagir com alvos
17.
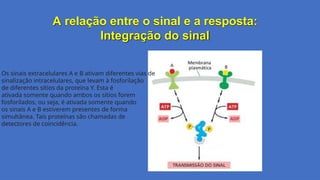
A relação entreo sinal e a resposta:
Integração do sinal
Os sinais extracelulares A e B ativam diferentes vias de
sinalização intracelulares, que levam à fosforilação
de diferentes sítios da proteína Y. Esta é
ativada somente quando ambos os sítios forem
fosforilados, ou seja, é ativada somente quando
os sinais A e B estiverem presentes de forma
simultânea. Tais proteínas são chamadas de
detectores de coincidência.
18.
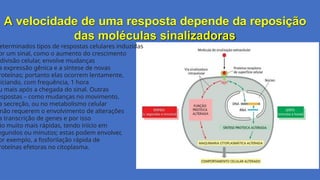
A velocidade deuma resposta depende da reposição
das moléculas sinalizadoras
eterminados tipos de respostas celulares induzidas
or um sinal, como o aumento do crescimento
divisão celular, envolve mudanças
a expressão gênica e a síntese de novas
roteínas; portanto elas ocorrem lentamente,
niciando, com frequência, 1 hora
u mais após a chegada do sinal. Outras
espostas – como mudanças no movimento,
a secreção, ou no metabolismo celular
não requerem o envolvimento de alterações
a transcrição de genes e por isso
ão muito mais rápidas, tendo início em
egundos ou minutos; estas podem envolver,
or exemplo, a fosforilação rápida de
roteínas efetoras no citoplasma.
19.
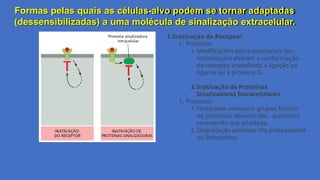
Formas pelas quaisas células-alvo podem se tornar adaptadas
(dessensibilizadas) a uma molécula de sinalização extracelular.
Mecanismos de dessensibilização celular,
são mecanismos pelos quais as células
reduzem ou interrompem sua resposta a
uma molécula de sinalização extracelular
após exposição prolongada ou repetida.
Esses processos evitam hiperatividade e
1.Sequestro do Receptor:
1. Processo:
1. Após a ativação, os receptores (ex.: GPCRs,
receptores tirosina-quinase)
são internalizados via endocitose mediada por
clatrina.
2. São transportados para endossomos e,
posteriormente, para lisossomos, onde são
degradados.
2. Efeito: Reduz o número de receptores disponíveis
na membrana plasmática, diminuindo a
sensibilidade celular ao sinal.
2.Retrorregulação do Receptor:
1. Processo:
1. Feedback negativo: A ativação do receptor
desencadeia vias que suprimem sua própria
atividade.
2. Efeito: Interrompe a sinalização mesmo na
presença contínua do ligante.
20.
Formas pelas quaisas células-alvo podem se tornar adaptadas
(dessensibilizadas) a uma molécula de sinalização extracelular.
1.Inativação do Receptor:
1. Processo:
1. Modificações pós-traducionais (ex.:
fosforilação) alteram a conformação
do receptor, impedindo a ligação ao
ligante ou à proteína G.
2.Inativação de Proteínas
Sinalizadoras Intracelulares:
1. Processo:
1. Fosfatases removem grupos fosfato
de proteínas ativadas (ex.: quinases),
revertendo sua atividade.
2. Degradação proteica: Via proteassoma
ou lisossomos.
21.
Formas pelas quaisas células-alvo podem se tornar adaptadas
(dessensibilizadas) a uma molécula de sinalização extracelular.
1.Produção de Proteínas Inibidoras:
1. Processo:
1. A célula sintetiza proteínas que bloqueiam
componentes da via de sinalização.
2. Exemplos:
1.Inibidores de quinases (ex.: PKI, que inibe a
PKA).
2.Proteínas sequestradoras (ex.: Gβγ, que
competem por alvos).
2. Efeito: Interrompe a transdução do sinal em múltiplos
níveis.
22.
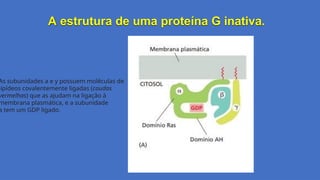
A estrutura deuma proteína G inativa.
As subunidades a e y possuem moléculas de
lipídeos covalentemente ligadas (caudas
vermelhas) que as ajudam na ligação à
membrana plasmática, e a subunidade
a tem um GDP ligado.
23.
Ativação de umaproteína G por um GPCR ativado
Passos da Ativação da Proteína G:
1. Uma molécula sinalizadora extracelular (ex.: hormônio,
neurotransmissor) liga-se ao domínio extracelular do GPCR,
localizado na membrana plasmática.
2. A ligação induz uma alteração estrutural no GPCR,
expondo um sítio de interação para a proteína G inativa
(ligada a GDP).
3. O GPCR ativado promove a substituição de GDP por
GTP na subunidade α da proteína G.
4. Essa troca é facilitada por fatores de troca de
nucleotídeos (GEFs).
5. A subunidade α (agora ligada a GTP) dissocia-se do
complexo βγ.
6. Tanto a subunidade α-GTP quanto o dímero βγ tornam-se
ativos e capazes de interagir com efetores.
7. Subunidade α-GTP: Ativa enzimas como a adenilil
ciclase (gera AMP cíclico) ou fosfolipase C (produz IP₃ e
DAG).
8. Dímero βγ: Pode modular canais iônicos ou outras vias de
sinalização (ex.: canais de K⁺).
24.
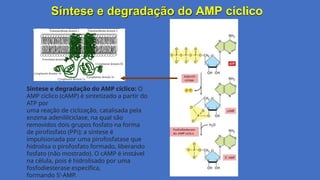
Síntese e degradaçãodo AMP cíclico
Síntese e degradação do AMP cíclico: O
AMP cíclico (cAMP) é sintetizado a partir do
ATP por
uma reação de ciclização, catalisada pela
enzima adenililciclase, na qual são
removidos dois grupos fosfato na forma
de pirofosfato (PPi); a síntese é
impulsionada por uma pirofosfatase que
hidrolisa o pirofosfato formado, liberando
fosfato (não mostrado). O cAMP é instável
na célula, pois é hidrolisado por uma
fosfodiesterase específica,
formando 5’-AMP.
26.
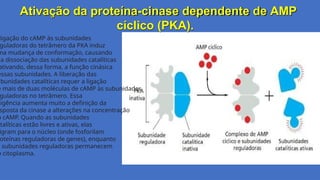
Ativação da proteína-cinasedependente de AMP
cíclico (PKA).
ligação do cAMP às subunidades
guladoras do tetrâmero da PKA induz
ma mudança de conformação, causando
ua dissociação das subunidades catalíticas
ativando, dessa forma, a função cinásica
essas subunidades. A liberação das
ubunidades catalíticas requer a ligação
e mais de duas moléculas de cAMP às subunidades
guladoras no tetrâmero. Essa
xigência aumenta muito a definição da
sposta da cinase a alterações na concentração
o cAMP. Quando as subunidades
atalíticas estão livres e ativas, elas
igram para o núcleo (onde fosforilam
roteínas reguladoras de genes), enquanto
s subunidades reguladoras permanecem
o citoplasma.
27.
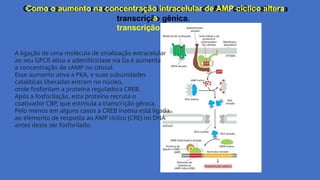
Como o aumentona concentração intracelular de AMP cíclico altera
a
transcrição gênica.
A ligação de uma molécula de sinalização extracelular
ao seu GPCR ativa a adenililciclase via Gs e aumenta
a concentração de cAMP no citosol.
Esse aumento ativa a PKA, e suas subunidades
catalíticas liberadas entram no núcleo,
onde fosforilam a proteína reguladora CREB.
Após a fosforilação, esta proteína recruta o
coativador CBP, que estimula a transcrição gênica.
Pelo menos em alguns casos a CREB inativa está ligada
ao elemento de resposta ao AMP cíclico (CRE) no DNA
antes deste ser fosforilado.
28.
Como o aumentona concentração intracelular de AMP cíclico altera
a
transcrição gênica.
Gláucia M1a9rques
Etapas da Sinalização cAMP Transcrição Gênica:
→
1. Uma molécula sinalizadora extracelular (ex.: hormônio ou
neurotransmissor) liga-se a um GPCR na membrana plasmática.
2. O receptor ativado estimula uma proteína G (ex.: Gαs), que ativa
a adenilil ciclase (correção de "Adentilicidade ativada").
3. A adenilil ciclase converte ATP em cAMP, aumentando sua
concentração intracelular.
4. O cAMP liga-se às subunidades reguladoras (R) da PKA, liberando
as subunidades catalíticas (C) ativas.
5. As subunidades C da PKA migram para o núcleo através dos poros
nucleares.
6. No núcleo, a PKA fosforila o CREB (cAMP response element-binding
protein) em um resíduo específico de serina (ex.: Ser133).
7. A fosforilação ativa o CREB, permitindo que ele se liga a elementos
de resposta ao cAMP (CRE) no DNA.
8. O CREB fosforilado interage com a CBP (CREB-binding protein), uma
proteína coativadora que possui atividade histona
acetiltransferase.
9. A CBP recruta componentes da máquina transcricional basal (ex.:
RNA polimerase II) e remodela a cromatina, facilitando o acesso ao
29.
A prpteína Gtransmite sinais através de fosfolipídeos
30.
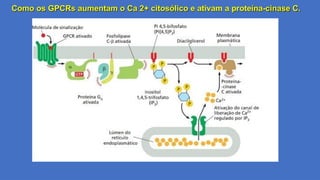
Como os GPCRsaumentam o Ca 2+ citosólico e ativam a proteína-cinase C.
31.
Como os GPCRsaumentam o Ca 2+ citosólico e ativam a proteína-cinase C.
Etapas da Sinalização
1. Uma molécula sinalizadora extracelular (ex.: acetilcolina, hormônios peptídicos) liga-se ao GPCR na
membrana plasmática.
2. O receptor ativado interage com uma proteína G do tipo Gq, que está ligada a GDP em seu estado
inativo.
3. O GPCR ativado atua como fator de troca de nucleotídeos (GEF), promovendo a substituição de GDP
por GTP na subunidade α da proteína Gq.
4. A subunidade α-GTP se dissocia do complexo βγ e ativa sua enzima efetora: a fosfolipase C-β (PLCβ).
5. A PLCβ cliva o fosfatidilinositol 4,5-bifosfato (PIP₂), um fosfolipídeo da membrana plasmática, em dois
segundos mensageiros: Inositol 1,4,5-trifosfato(IP3) e Diacilglicerol (DAG)
6. O IP₃ liga-se a receptores de IP₃ (IP₃R) no RE, que funcionam como canais de cálcio.
7. A abertura desses canais libera Ca²⁺ armazenado no lúmen do RE para o citosol, elevando sua
concentração citosólica.
8. O DAG na membrana e o Ca²⁺ citosólico atuam em conjunto para recrutar e ativar a PKC.
9. A PKC transloca-se para a membrana plasmática, onde:
1. DAG a ancora na membrana.
2. Ca²⁺ induz uma mudança conformacional que expõe o sítio catalítico da enzima.
10.A PKC ativada fosforila proteínas-alvo em resíduos de serina/treonina, desencadeando respostas
celulares.
32.
Papel do óxidonítrico (NO) no relaxamento do músculo liso da
parede de um vaso sanguíneo
32
(A) Corte transversal
simplificado de um vaso
sanguíneo, mostrando as
células endoteliais revestindo
o lúmen e as células
musculares lisas em volta
delas.
33.
33
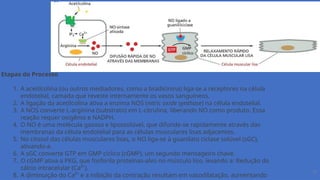
Etapas do Processo
1.A acetilcolina (ou outros mediadores, como a bradicinina) liga-se a receptores na célula
endotelial, camada que reveste internamente os vasos sanguíneos.
2. A ligação da acetilcolina ativa a enzima NOS (nitric oxide synthase) na célula endotelial.
3. A NOS converte L-arginina (substrato) em L-citrulina, liberando NO como produto. Essa
reação requer oxigênio e NADPH.
4. O NO é uma molécula gasosa e lipossolúvel, que difunde-se rapidamente através das
membranas da célula endotelial para as células musculares lisas adjacentes.
5. No citosol das células musculares lisas, o NO liga-se à guanilato ciclase solúvel (sGC),
ativando-a.
6. A sGC converte GTP em GMP cíclico (cGMP), um segundo mensageiro chave.
7. O cGMP ativa a PKG, que fosforila proteínas-alvo no músculo liso, levando a: Redução do
cálcio intracelular (Ca²⁺).
8. A diminuição do Ca²⁺ e a inibição da contração resultam em vasodilatação, aumentando
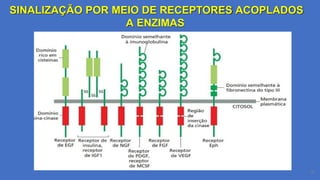
SINALIZAÇÃO POR MEIODE RECEPTORES ACOPLADOS
A ENZIMAS
36
Componentes Estruturais dos RTKs:
1.Domínios Extracelulares:
1. Variam entre os diferentes tipos de RTKs e são responsáveis pelo reconhecimento específico de ligantes.
Exemplos:
1. Domínio semelhante à imunoglobulina: Encontrado em receptores como o EGFR (Epidermal Growth
Factor Receptor), permite interação com fatores de crescimento.
2. Domínio rico em cisteínas: Presente no receptor de insulina, estabiliza a ligação ao hormônio.
3. Domínio semelhante à fibronectina tipo III: Auxilia na interação com a matriz extracelular
(ex.: receptor de FGF).
4. Domínio de ligação a efrina: Específico para receptores Eph, envolvidos em orientação celular
durante o desenvolvimento.
2.Domínio Transmembrana:
1. Hélice α hidrofóbica: Ancora o receptor na membrana plasmática, conectando os domínios extracelular e
intracelular.
3.Domínio Intracelular:
1. Região da tirosina quinase: Contém atividade enzimática para autofosforilação e fosforilação de
substratos.