Baixado 805 vezes























































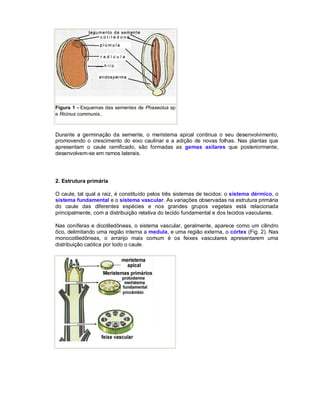


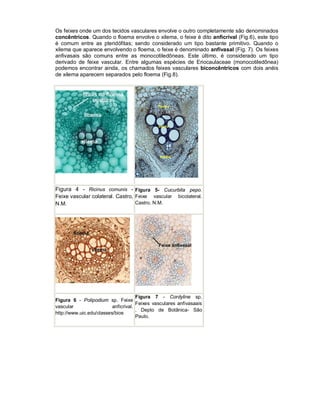







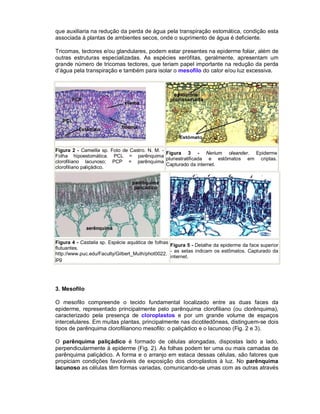
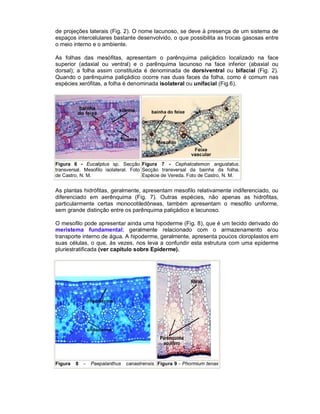











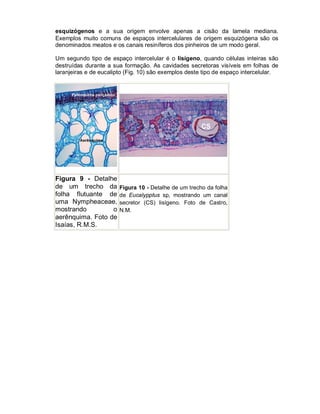

Este documento descreve os meristemas em plantas. Os meristemas são tecidos que permanecem embrionários e conservam a capacidade de divisão celular, responsáveis pelo crescimento das plantas. Existem três tipos principais de meristemas: apicais, localizados nos ápices de raízes e caules; laterais, como o câmbio vascular e o felogênio; e intercalares, entre tecidos maduros. Os meristemas apicais formam a estrutura primária da planta e os laterais formam a estrutura sec


























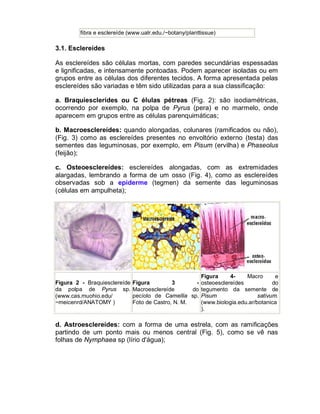
![Aula Biologia: introdução à biologia [1° Ano Ensino Médio]](https://cdn.slidesharecdn.com/ss_thumbnails/mdulo1-introduobiologia-170129000526-thumbnail.jpg?width=640&height=640&fit=bounds)








































