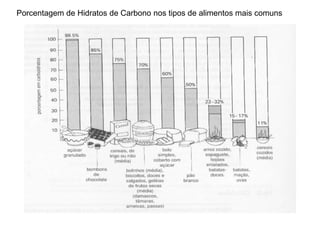
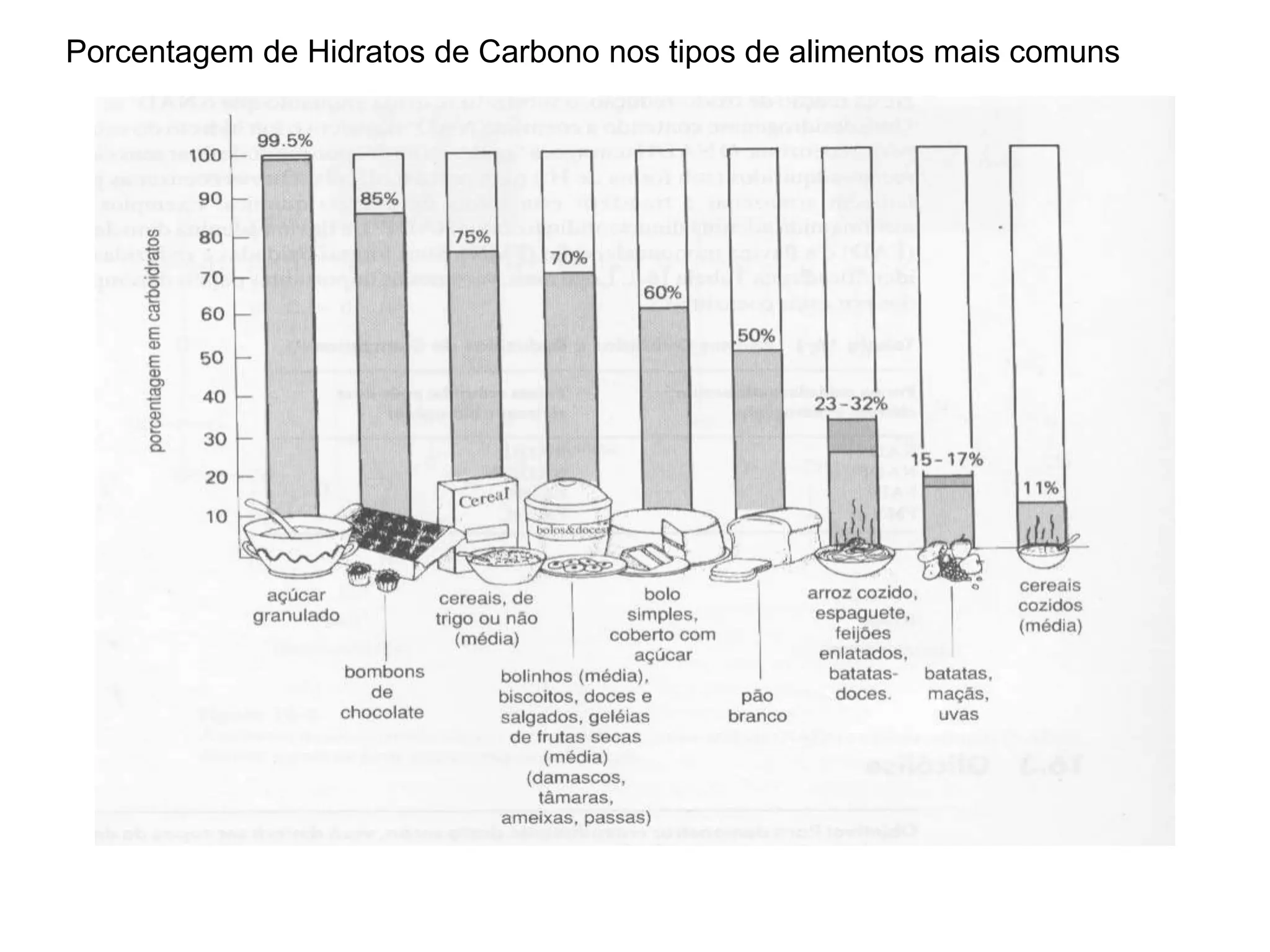
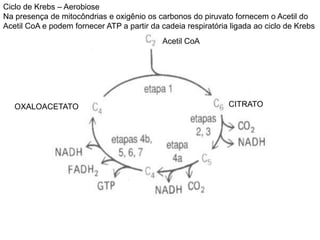
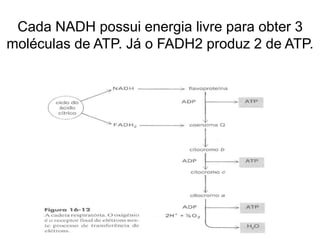
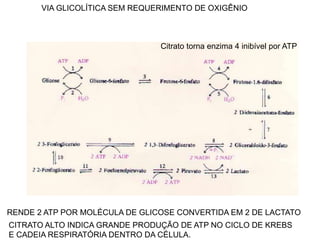
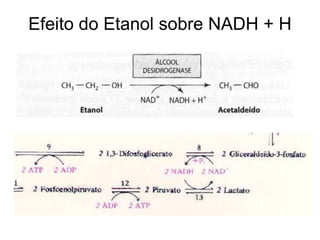
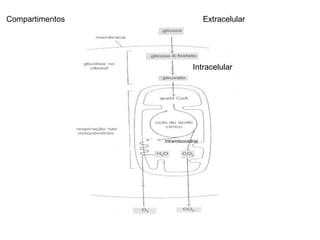
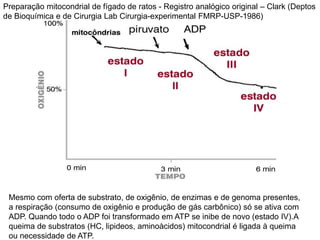
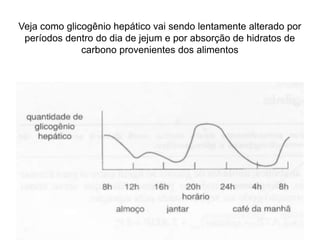
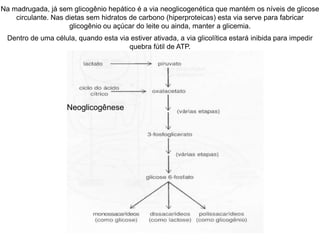
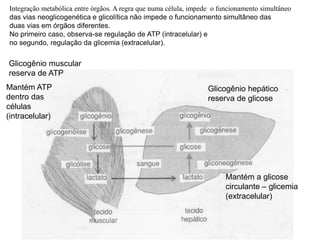
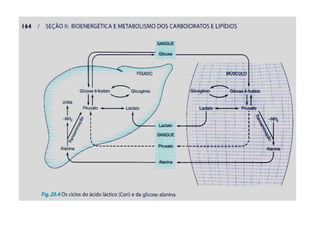
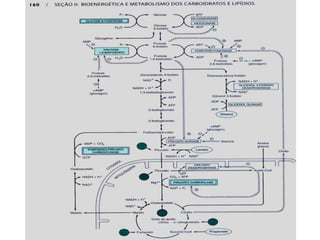
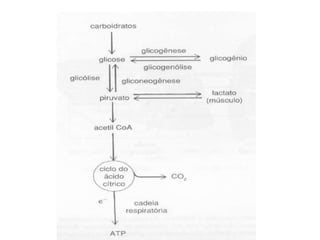
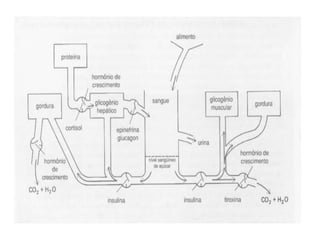
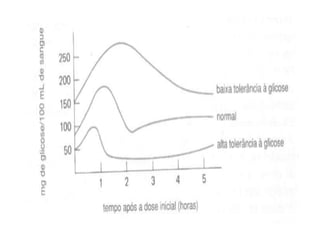
O documento discute a porcentagem de hidratos de carbonos nos principais tipos de alimentos e o metabolismo desses nutrientes no corpo humano. Aborda os processos de glicólise, ciclo de Krebs e cadeia respiratória na produção de energia a partir de glicose e piruvato, bem como a integração desses processos entre os diferentes órgãos. Também menciona a neoglicogênese hepática e sua importância para manter os níveis de glicose no sangue durante o jejum.