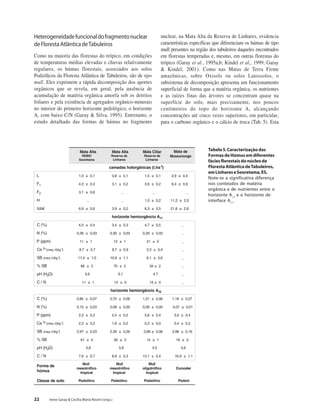
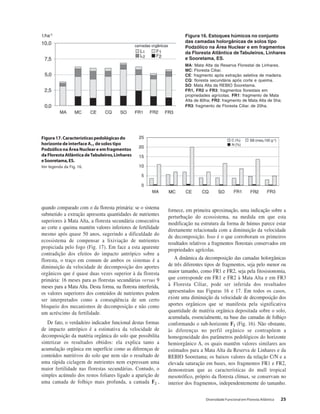
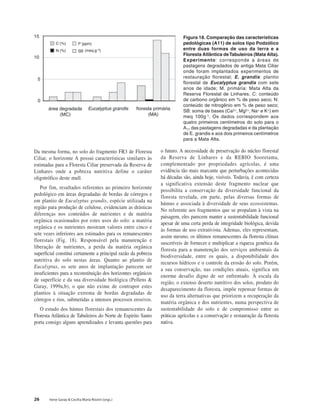
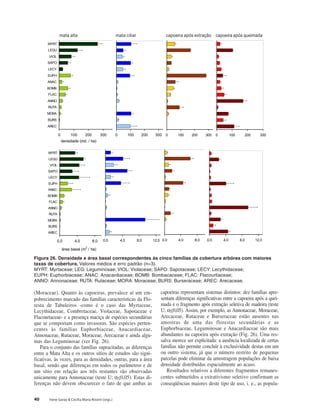
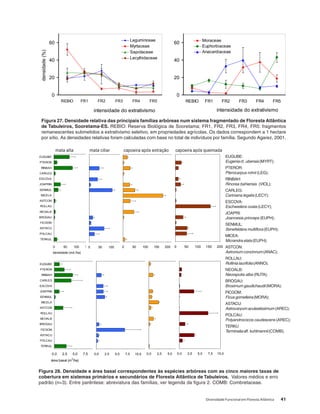
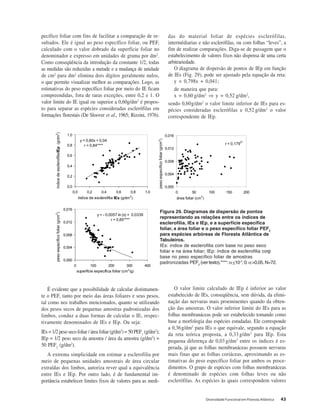
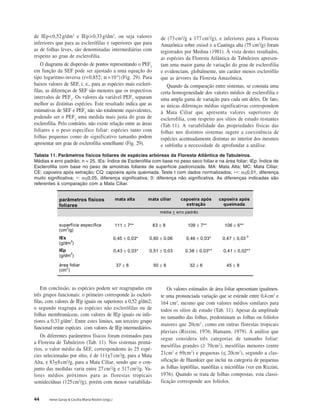
Baixar para ler offline

































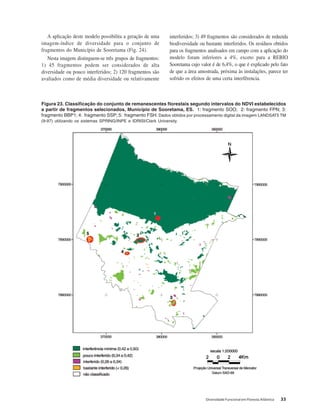
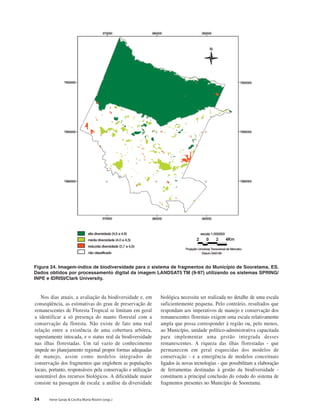



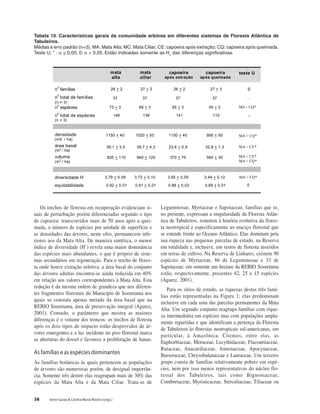











This document discusses the functional diversity of tree cover in the Atlantic Forest of Tabuleiro regions in Brazil. It focuses on the physical geography, land use, soil diversity, fragmented forest conservation, and leaf traits like sclerophylly that indicate biodiversity status across this forest type.




































































