Baixar para ler offline




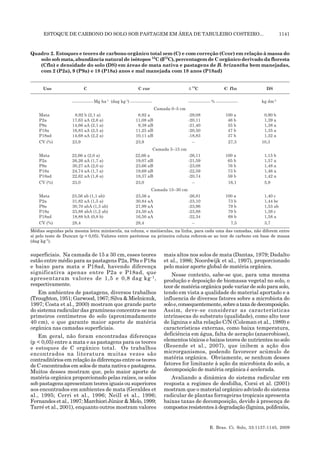





Este estudo avaliou o estoque de carbono no solo em áreas de pastagem degradada e produtiva com diferentes idades de uso, comparando-as com o solo de uma mata natural, em Itabela, Bahia. Os resultados mostraram que não houve diferença significativa nos estoques de carbono do solo entre a mata, pasto degradado e pasto produtivo nas diferentes camadas de solo. Além disso, observou-se que após 28 anos de uso, em média 62% do carbono orgânico no solo de pastagens bem ou mal manejadas ainda era derivado





















































