Genética Geral
Para Universitários
PauloRoberto Eleutério de Souza
Hildson Dornelas Angelo da Silva
Fernanda Cristina Bezerra Leite
Maria de Mascena Diniz Maia
Ana Cristina Lauer Garcia
Martín Alejandro Montes
2.
Genética Geral
Para Universitários
PauloRoberto Eleutério de Souza
Hildson Dornelas Angelo da Silva
Fernanda Cristina Bezerra Leite
Maria de Mascena Diniz Maia
Ana Cristina Lauer Garcia
Martín Alejandro Montes
Recife | 2015
1a Edição
3.
Profª. Maria Joséde Sena — Reitora
Prof. Marcelo Brito Carneiro Leão — Vice-Reitor
Conselho Editorial
Presidente: Marcelo Brito Carneiro Leão; Diretor da Editora da UFRPE: Bruno de Souza
Leão; Diretora do Sistema de Bibliotecas da UFRPE : Maria Wellita Santos
Conselheiros: Álvaro José de Almeida Bicudo, Fernando Joaquim Ferreira Maia
Maria do Rosário de Fátima Andrade, Monica Lopes Folena Araújo, Rafael Miranda
Tassitano, Renata Pimentel Teixeira
Revisão final dos autores
Editora filiada à
Positivo e negativo
Manual de Identidade Visual ABEU 07
Todos os direitos reservados. É proibida a reprodução total ou parcial do trabalho sem autorização da
universidade federal rural de pernambuco - departamento de biologia - genética e dos autores.
Ficha catalográfica
G328 Genética geral para universitários / Maria de Mascena Diniz
Maia, coordenadora; Paulo Roberto Eleutério de Souza ...
[et al]. -- 1. ed. -- Recife : EDUFRPE, 2015.
147 p. : il.
ISBN : 978-85-7946-229-0
Referências.
1. Genética I. Maia, Maria de Mascena Diniz, coord.
II. Souza, Paulo Roberto Eleutério de
CDD 575.1
4.
DEDICATÓRIAS
Aos meus filhos,Paula, Bruno e Alexandre
e minhas netas, Maria Gabriela e Maria Luísa.
Maria de Mascena Diniz Maia
À Katarina, meu complemento, à Marinho,
mestre plenamente abençoado por Deus e à
minha família.
Hildson Dornelas
A nossa F1 (Mariana).
Ana Cristina Lauer Garcia
e Martín Alejandro Montes
Aos meus pais, Severino e Betânia, meu
esposo, George e minha filha, Maria Fernanda.
Fernanda Cristina Bezerra Leite
A minha esposa Fabianne
e meu filho Felipe.
Paulo Roberto Eleutério Souza
6.
AGRADECIMENTOS
A Reitoria daUFRPE pelo apoio na confecção
deste exemplar, a Editora Universitária da UFRPE
pela Editoração, impressão e encadernação, aos
Professores autores dos capítulos deste livro e
um especial agradecimento ao Professor Hild-
son Dornelas pela contribuição na organização
deste exemplar.
8.
PREFÁCIO
Desde ano de2008, vimos programando editar um livro de genética geral,
para ser usado pelos alunos dos cursos de Agronomia, Engenharia Florestal,
Veterinária, Zootecnia, Biologia (Licenciatura e Bacharelado) da UFRPE. A
intenção foi descrever os temas do conteúdo da disciplina Genética Geral, em
uma linguagem simples, lógica e sintética com o objetivo de facilitar o aluno
na compreensão dos conceitos básicos de cada assunto abordado pertinente
ao processo de aprendizagem da genética. Essa ideia nasceu da experiência
como professores de genética sempre ouvindo dos alunos que essa disciplina
é “muito difícil”, os “livros trazem uma linguagem complexa” os “assuntos
envolvem matemática” os “capítulos são longos” entre outras dificuldades.
Assim, em 2011, resolvemos elaborar um livro para ser publicado pela
Editora da Universidade Federal Rural de Pernambuco. Desejamos, portanto,
que os nossos alunos em contato com os assuntos que escrevemos sejam esti-
mulados a aprender genética de uma forma pessoal relacionando os conteúdos
a algo que eles já conhecem e se deparam no dia-a-dia associando os fatos
do passado com o presente de modo a tornar os temas mais interessantes no
que se refere à beleza sobre o panorama da vida.
10.
SUMÁRIO
13 CICLO CELULAR
13Introdução
13 Intérfase
14 Mitose
18 Meiose
21 Controle do Ciclo Celular
21 Substâncias Indutoras
23 Supressores Tumorais
24 Exercícios
31 PRINCÍOS BÁSICOS DE HEREDITARIEDADE
31 O que é genética?
31 Ideias sobre herança antes de Mendel
32 Quem foi Mendel
33 A primeira Lei de Mendel
39 A primeira Lei de Mendel hoje
41 Cruzamento teste
42 A segunda Lei de Mendel
45 A segunda Lei de Mendel Hoje
45 Diagrama ramificado
48 Exercícios
51 VARIAÇÃO ALÉLICA E FUNCIONAMENTO GÊNICO
51 Variação alélica e funcionamento gênico
51 Dominância incompleta
52 Codominância
53 Alelos Múltiplos
56 Por que Algumas Mutações
56 são Dominantes e Outras Recessivas?
58 Influência do Ambiente na Expressão do Gene
59 Penetrância e Expressividade
59 Interações Gênicas
11.
60 Interação Gênicanão Epistática
61 Interação Gênica Epistática
61 Características da interação gênica epistática:
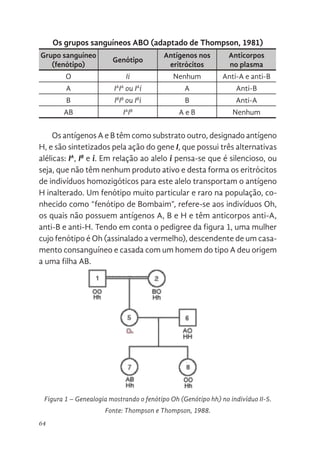
63 Interação Gênica Epistática em Humanos
67 Exercícios
75 MAPEAMENTO CROMOSSÔMICO
75 Introdução
77 Princípio da distribuição independente
78 Teoria Cromossômica da Hereditariedade
79 Experimento de Bridges para comprovar que o gene para olhos brancos da Droso-
phila está situado no cromossomo X (Teoria Cromossômica da Herança).
80 Ligação, Recombinação e Crossing over
83 Ligação e Recombinação entre dois genes
84 Cálculo da Frequência de Recombinação
86 Mapeamento gênico em eucarioto
87 Recombinação com cruzamento-teste de dois pontos
89 Mapeamento de recombinação
89 em cruzamento-teste de três pontos
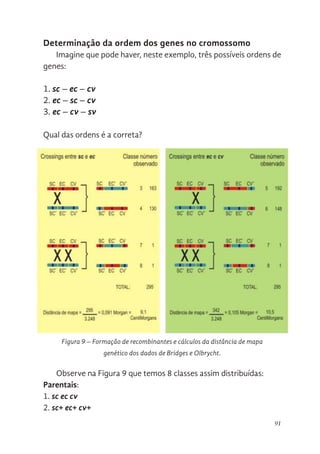
91 Determinação da ordem dos genes no cromossomo
92 Cálculo das distâncias entre os genes
94 DETERMINAÇÃO DO SEXO
96 Mecanismos de Determinação do Sexo
96 Sistemas de Determinação Ambiental do Sexo
98 Sistema Haploide/Diploide de Determinação do Sexo
98 Sistema de Determinação do Sexo pela Relação entre
Cromossomos Sexuais e Não Sexuais
99 Sistemas de Determinação Cromossômica do Sexo
101 Determinação do Sexo em Humanos
102 Exercícios
105 ALTERAÇÕES CROMOSSÔMICAS
105 Cromossomos: composição e estrutura
105 Cromossomos metafásicos e o cariótipo
12.
106 Alterações cromossômicas:Numéricas e Estruturais
107 Alterações Numéricas
109 Alterações Estruturais
112 Exercícios
114 ESTRUTURA E REPLICAÇÃO DO DNA
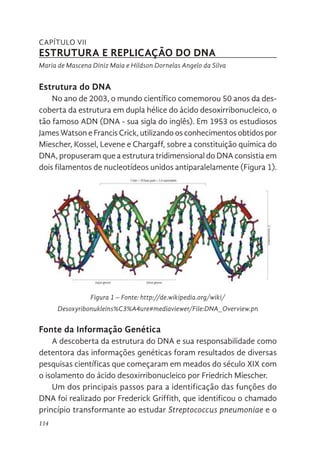
114 Estrutura do DNA
114 Fonte da Informação Genética
116 Funções do Material Genético
117 Estrutura Primária
117 Componentes dos Nucleotídeos
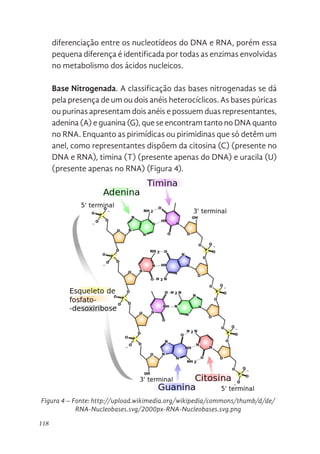
119 Complementariedade das Bases Nitrogenadas
119 DNA é uma hélice
119 A “Descoberta” da Dupla Hélice
120 Estrutura Secundária
120 Formas Alternativas da Dupla Hélice
120 Replicação do Dna
121 Replicação Semiconservativa
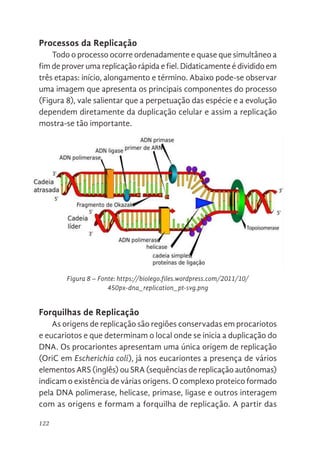
122 Processos da Replicação
122 Forquilhas de Replicação
123 Separação das fitas e giro
123 Estabilização da desnaturação
123 Iniciação da replicação
124 Polimerização
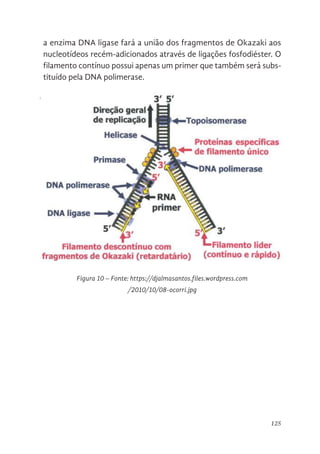
124 Fragmento de Okazaki e DNA ligase
126 Exercícios
132 TRANSCRIÇÃO E TRADUÇÃO
132 Transcrição
132 Classes de RNA
134 Sequências promotoras
134 RNA polimerase
136 Etapas da transcrição
136 Iniciação da transcrição
137 Alongamento da Transcrição
13.
138 Término daCadeia de RNA
138 Processamento do RNA
138 Capeamento do RNA mensageiro
139 Poliadenilação do RNA mensageiro
139 Edição do RNA
139 Splicing
140 Tradução
142 Etapas da Síntese Protéica
142 Inicio
143 Alongamento
144 Término
146 Exercícios
149 MUTAÇÃO E REPARO DO DNA
149 Mutações Gênicas: Definição e Importância
149 Como Surgem as Mutações?
153 Onde Ocorrem as Mutações?
153 Como se Classificam as Mutações?
156 Mecanismos de Reparo
157 Reparo de Pareamento Errado
157 Reparo Direto
157 Reparo por Excisão de Bases
160 Reparo por Excisão de Nucleotídeos
161 Exercícios
165 REFERÊNCIAS BIBLIOGRÁFICAS
166 INFORMAÇÕES DOS AUTORES
14.
13
CAPÍTULO I
CICLO CELULAR
PauloRoberto Eleutério de Souza e Hildson Dornelas Angelo da Silva
Introdução
O ciclo celular corresponde ao mecanismo de divisão de células
de origem somática e de origem da linhagem germinativa. Este me-
canismo é fundamental para originar tecidos, órgãos e os gametas.
Este mecanismo é regulado por um grupo de proteínas que sinalizam
a passagem de um estágio para outro do ciclo celular. Alterações
no mecanismo de regulação resultam na perda do controle do ciclo
celular acarretando numa multiplicação desordenada podendo gerar
o desenvolvimento de células anormais, tendo como consequência o
desenvolvimento de câncer.
Intérfase
A divisão celular é precedida da Interfase que é um período de
intensa atividade metabólica necessária para preparar a célula para
a divisão celular propriamente dita. A interfase é dividida em três
estágios: G1 (do inglês, gap, intervalo 1), S (de síntese do DNA) e G2
(do inglês, gap, intervalo 2) .
No estágio G1, ocorre intensa síntese de moléculas de RNA que
vão para o citoplasma, aumento do volume celular e produção de
proteínas que serão utilizadas no processo de multiplicação do mate-
rial genético celular. Células permanentes (como as nervosas ou as do
músculo esquelético) permanecem estacionadas no período G1 por
toda a vida, por isso costuma-se denominar que se encontra em G0
.
A duração deste período é geralmente de 4 horas. Antes de a célula
entrar na fase S, é necessário ultrapassar o ponto crítico de controle do
ciclo celular ou ponto de verificação, chamado de G1/S, que garante
que sejam sintetizadas todas as proteínas necessárias a replicação do
DNA. Sinais regulatórios podem manter as células sem divisão celular,
ou seja, as células são mantidas no estágio G0
por um período.
15.
14
No estágio S,ocorre a duplicação dos cromossomos de forma
semiconservativa. É o período mais crítico e importante do ciclo celu-
lar. No final deste estágio cada cromossomo será composto por duas
cromátides exatamente iguais (cromátides irmãs).
E no estágio G2, ocorre aumento do volume celular e síntese de
proteínas que serão utilizadas na divisão celular propriamente dita.
Nesta fase, ocorrem vários eventos bioquímicos essenciais a divisão
celular e um ponto adicional de controle do estágio G2 para mitose,
chamado ponto crítico G2/M, para evitar que a multiplicação celular
ocorra de forma alterada. Enzimas fazem a revisão das sequências de
nucleotídeos do material genético e caso encontrem erros, tentam
repará-los. Caso os erros persistam as células normalmente encami-
nham-se para o processo de apoptose (morte programada), como
ação preventiva, para que células defeituosas não continuem vivas e
possam se multiplicar podendo gerar principalmente câncer.
Mitose
A mitose é o processo de divisão celular que é característico
de células da linhagem somática (ressalta-se a mitose em gônias,
células da linhagem germinativa) e é responsável pela formação de
todos os órgãos e tecidos de um indivíduo. A mitose é precedida da
interfase, explicada acima. Na mitose o tipo de divisão celular é dita
equacional, pois mantém o número de cromossomos da célula mãe
(origem). A partir de uma célula obtêm-se duas células exatamente
iguais. O processo ocorre em quatro fases a saber: Prófase, Metáfase,
Anáfase e Telófase.
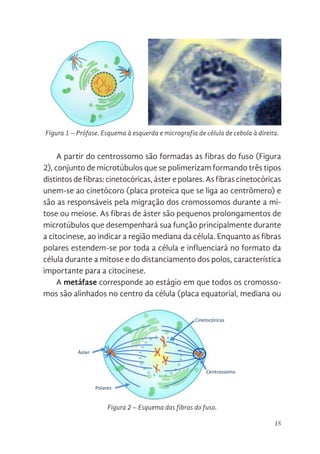
A prófase é o primeiro estágio da mitose (Figura 1). É carac-
terizada pela desintegração da carioteca (membrana que recobre
o núcleo celular), desaparecimento dos nucléolos, condensação
da cromatina, duplicação e migração dos centrossomos para os
polos opostos (estrutura que contém os centríolos), formação das
fibras do fuso e deslocamento dos cromossomos para a região
central da célula.
16.
15
A partir docentrossomo são formadas as fibras do fuso (Figura
2), conjunto de microtúbulos que se polimerizam formando três tipos
distintos de fibras: cinetocóricas, áster e polares. As fibras cinetocóricas
unem-se ao cinetócoro (placa proteica que se liga ao centrômero) e
são as responsáveis pela migração dos cromossomos durante a mi-
tose ou meiose. As fibras de áster são pequenos prolongamentos de
microtúbulos que desempenhará sua função principalmente durante
a citocinese, ao indicar a região mediana da célula. Enquanto as fibras
polares estendem-se por toda a célula e influenciará no formato da
célula durante a mitose e do distanciamento dos polos, característica
importante para a citocinese.
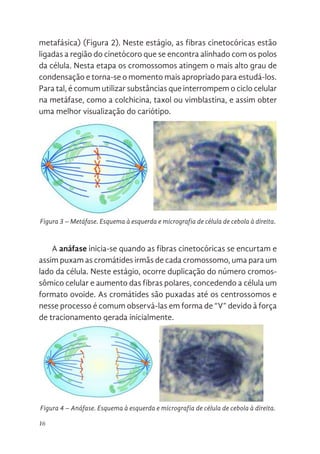
A metáfase corresponde ao estágio em que todos os cromosso-
mos são alinhados no centro da célula (placa equatorial, mediana ou
12
A partir do centrossomo são formadas as fibras do fuso (Figura 2),
conjunto de microtúbulos que se polimerizam formando três tipos distintos de
fibras: cinetocóricas, áster e polares. As fibras cinetocóricas unem-se ao
cinetócoro (placa proteica que se liga ao centrômero) e são as responsáveis
pela migração dos cromossomos durante a mitose ou meiose. As fibras de
áster são pequenos prolongamentos de microtúbulos que desempenhará sua
função principalmente durante a citocinese, ao indicar a região mediana da
célula. Enquanto as fibras polares estendem-se por toda a célula e influenciará
no formato da célula durante a mitose e do distanciamento dos polos,
característica importante para a citocinese.
Áster
Polares
Cinetocóricas
Centrossomo
Figura 2. Esquema das fibras do fuso.
Figura 1. Prófase. Esquema à esquerda e micrografia de célula de cebola à
direita.
12
A partir do centrossomo são formadas as fibras do fuso (Figura 2),
conjunto de microtúbulos que se polimerizam formando três tipos distintos de
fibras: cinetocóricas, áster e polares. As fibras cinetocóricas unem-se ao
cinetócoro (placa proteica que se liga ao centrômero) e são as responsáveis
pela migração dos cromossomos durante a mitose ou meiose. As fibras de
áster são pequenos prolongamentos de microtúbulos que desempenhará sua
função principalmente durante a citocinese, ao indicar a região mediana da
célula. Enquanto as fibras polares estendem-se por toda a célula e influenciará
no formato da célula durante a mitose e do distanciamento dos polos,
característica importante para a citocinese.
Áster
Polares
Cinetocóricas
Centrossomo
Figura 2. Esquema das fibras do fuso.
Figura 1. Prófase. Esquema à esquerda e micrografia de célula de cebola à
direita.
Figura 1 – Prófase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 2 – Esquema das fibras do fuso.
17.
16
metafásica) (Figura 2).Neste estágio, as fibras cinetocóricas estão
ligadas a região do cinetócoro que se encontra alinhado com os polos
da célula. Nesta etapa os cromossomos atingem o mais alto grau de
condensação e torna-se o momento mais apropriado para estudá-los.
Para tal, é comum utilizar substâncias que interrompem o ciclo celular
na metáfase, como a colchicina, taxol ou vimblastina, e assim obter
uma melhor visualização do cariótipo.
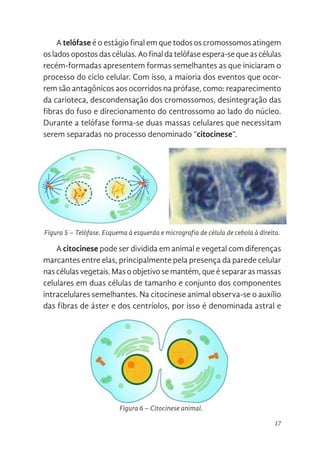
A anáfase inicia-se quando as fibras cinetocóricas se encurtam e
assim puxam as cromátides irmãs de cada cromossomo, uma para um
lado da célula. Neste estágio, ocorre duplicação do número cromos-
sômico celular e aumento das fibras polares, concedendo a célula um
formato ovoide. As cromátides são puxadas até os centrossomos e
nesse processo é comum observá-las em forma de “V” devido à força
de tracionamento gerada inicialmente.
13
A metáfase corresponde ao estágio em que todos os cromossomos são
alinhados no centro da célula (placa equatorial, mediana ou metafásica) (Figura
2). Neste estágio, as fibras cinetocóricas estão ligadas a região do cinetócoro
que se encontra alinhado com os polos da célula. Nesta etapa os
cromossomos atingem o mais alto grau de condensação e torna-se o momento
mais apropriado para estudá-los. Para tal, é comum utilizar substâncias que
interrompem o ciclo celular na metáfase, como a colchicina, taxol ou
vimblastina, e assim obter uma melhor visualização do cariótipo.
A anáfase inicia-se quando as fibras cinetocóricas se encurtam e assim
puxam as cromátides irmãs de cada cromossomo, uma para um lado da célula.
Neste estágio, ocorre duplicação do número cromossômico celular e aumento
das fibras polares, concedendo a célula um formato ovoide. As cromátides são
puxadas até os centrossomos e nesse processo é comum observá-las em
forma de “V” devido à força de tracionamento gerada inicialmente.
Figura 3. Metáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
13
A metáfase corresponde ao estágio em que todos os cromossomos são
alinhados no centro da célula (placa equatorial, mediana ou metafásica) (Figura
2). Neste estágio, as fibras cinetocóricas estão ligadas a região do cinetócoro
que se encontra alinhado com os polos da célula. Nesta etapa os
cromossomos atingem o mais alto grau de condensação e torna-se o momento
mais apropriado para estudá-los. Para tal, é comum utilizar substâncias que
interrompem o ciclo celular na metáfase, como a colchicina, taxol ou
vimblastina, e assim obter uma melhor visualização do cariótipo.
A anáfase inicia-se quando as fibras cinetocóricas se encurtam e assim
puxam as cromátides irmãs de cada cromossomo, uma para um lado da célula.
Neste estágio, ocorre duplicação do número cromossômico celular e aumento
das fibras polares, concedendo a célula um formato ovoide. As cromátides são
puxadas até os centrossomos e nesse processo é comum observá-las em
forma de “V” devido à força de tracionamento gerada inicialmente.
Figura 3. Metáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 3 – Metáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 4. Anáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 4. Anáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 4 – Anáfase. Esquema à esquerda e micrografia de célula de cebola à direita.
18.
17
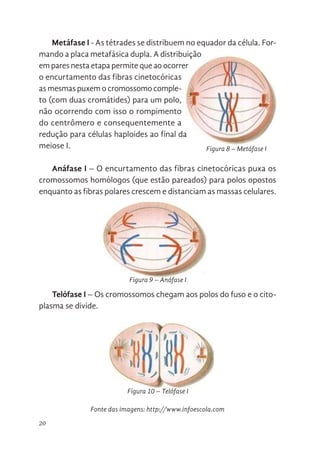
A telófase éo estágio final em que todos os cromossomos atingem
os lados opostos das células. Ao final da telófase espera-se que as células
recém-formadas apresentem formas semelhantes as que iniciaram o
processo do ciclo celular. Com isso, a maioria dos eventos que ocor-
rem são antagônicos aos ocorridos na prófase, como: reaparecimento
da carioteca, descondensação dos cromossomos, desintegração das
fibras do fuso e direcionamento do centrossomo ao lado do núcleo.
Durante a telófase forma-se duas massas celulares que necessitam
serem separadas no processo denominado “citocinese”.
A citocinese pode ser dividida em animal e vegetal com diferenças
marcantes entre elas, principalmente pela presença da parede celular
nas células vegetais. Mas o objetivo se mantém, que é separar as massas
celulares em duas células de tamanho e conjunto dos componentes
intracelulares semelhantes. Na citocinese animal observa-se o auxílio
das fibras de áster e dos centríolos, por isso é denominada astral e
14
A telófase é o estágio final em que todos os cromossomos atingem os
lados opostos das células. Ao final da telófase espera-se que as células recém-
formadas apresentem formas semelhantes as que iniciaram o processo do ciclo
celular. Com isso, a maioria dos eventos que ocorrem são antagônicos aos
ocorridos na prófase, como: reaparecimento da carioteca, descondensação dos
cromossomos, desintegração das fibras do fuso e direcionamento do
centrossomo ao lado do núcleo. Durante a telófase forma-se duas massas
celulares que necessitam serem separadas no processo denominado
“citocinese”.
Figura 5. Telófase. Esquema à esquerda e micrografia de célula de cebola à direita.
Figura 5 – Telófase. Esquema à esquerda e micrografia de célula de cebola à direita.
A citocinese pode ser dividida em animal e vegetal com diferenças
arcantes entre elas, principalmente pela presença da parede celular nas
lulas vegetais. Mas o objetivo se mantém, que é separar as massas celulares
m duas células de tamanho e conjunto dos componentes intracelulares
melhantes. Na citocinese animal observa-se o auxílio das fibras de áster e
s centríolos, por isso é denominada astral e cêntrica, respectivamente.
quanto há ausência de ambos nas células vegetais (anastral e acêntrica).
utra diferença está na forma de separação, nas células animais existe um
el contrátil formado de actina e miosina II que as estrangula no sentido extra
ra intracelular (citocinese centrípeta) (Figura 6). Já nas vegetais, os
lgiossomos produzem vesículas ricas em pectinas que se dispõem na placa
ediana das células e formam uma “parede” (fragmoplasto) de dentro para fora
tocinese centrífuga) que divide as duas células geradas.
eiose
Figura 6. Citocinese animal.
Figura 6 – Citocinese animal.
19.
18
cêntrica, respectivamente. Enquantohá ausência de ambos nas
células vegetais (anastral e acêntrica). Outra diferença está na
forma de separação, nas células animais existe um anel contrátil
formado de actina e miosina II que as estrangula no sentido extra
para intracelular (citocinese centrípeta) (Figura 6). Já nas vege-
tais, os golgiossomos produzem vesículas ricas em pectinas que
se dispõem na placa mediana das células e formam uma “parede”
(fragmoplasto) de dentro para fora (citocinese centrífuga) que
divide as duas células geradas.
Meiose
Para que ocorra a perpetuação das espécies o processo da
reprodução sexuada é praticamente obrigatório, já que, possibi-
lita uma mistura de material genético e assim garante uma maior
variabilidade genética. A ação de combinação entre materiais
genéticos dos parceiros sexuais se dá através da fecundação dos
gametas que são células modificadas que apresentam metade do
DNA de uma célula normal (diploide) e foram geradas através do
processo meiótico. Durante a formação do gameta, ocorreram
inúmeras trocas de segmentos entre cromossomos homólogos
(um paterno e outro materno), resultando em células haploides
únicas devido sua variabilidade genética, característica central do
processo meiótico.
A meiose é antecedida pela intérfase como a mitose, porém
ocorre em duas etapas (meiose I e meiose II) e é característica de
células germinativas (envolvidas na produção de gametas). A meio-
se I é denominada reducional, pois, ao final da mesma o número
cromossômico é reduzido pela metade, passando a ser haploide.
Enquanto a meiose II é conhecida como equacional, pois nela o nú-
mero cromossômico se mantém mesmo o material genético sendo
dividido entre as duas células filhas. Outra diferença marcante da
mitose é a quantidade de células geradas, ao final das 2 meioses,
observa-se 4 células filhas, enquanto apenas 2 na mitose.
20.
19
Paquíteno => Oscromossomos tornam-se mais condensad
grossos, e desenvolve-se um complexo sinaptonêmico entre os
homólogos. Ocorre à troca de segmentos entre cromossomo
denominada como permutação (crossing-over).
Diplóteno => Ocorre separação dos homólogos, mas os mesmos
ligados por pontos onde ocorreram os crossing overs, chamdos qu
Diacinese => A condensação cromossômica continua, e os quias
se para as pontas dos cromossomos, ocorrendo a termin
quiasmas.
Metáfase I - As tétrades se distribuem no equador da célu
a placa metafásica dupla. A distribuição em pares nesta etapa p
ocorrer o encurtamento das fibras cinetocóricas as mesm
Meiose I. Esta etapa é dividida em 4 fases: prófase I (Figura 7),
metáfase I (Figura 8), anáfase I (Figura 9) e telófase I (Figura 10).
Dentre elas, a prófase I é a mais extensa e importante, pois nela
ocorre a permutação ou crossing-over (no inglês), que é a troca de
segmentos de DNA entre cromossomos homólogos, permitindo com
isso a variabilidade genética pretendida pela meiose.
Prófase I - Os cromossomos condensam-se e os homólogos se
juntam formando tétrades; a carioteca e os nucléolos se desintegram;
os centrossomos duplicam e dirigem-se para os polos da célula; for-
mam-se as fibras do fuso. A prófase I é dividida em 5 subfases listadas
abaixo com seus principais acontecimentos:
Leptóteno → Aparecimento de pontos de rápida condensação, co-
nhecidos como, cromômeros. Neste estágio os cromossomos se
contraem e tornam-se visíveis.
Zigóteno → Os cromossomos homólogos ficam lado a lado, carac-
terizando a sinapse cromossômica. Cada par de homólogos em
sinapse forma tétrades (quatro cromátides juntas) ou bivalentes.
Os cromossomos continuam se condensando.
Paquíteno → Os cromossomos tornam-se mais condensados, curtos
e grossos, e desenvolve-se um complexo sinaptonêmico entre
os cromossomos homólogos. Ocorre à troca de segmentos
entre cromossomos homólogos, denominada como permutação
(crossing-over).
Diplóteno → Ocorre separação dos homó-
logos, mas os mesmos permanecem
ligados por pontos onde ocorreram os
crossing overs, chamdos quiasmas.
Diacinese → A condensação cromossômica
continua, e os quiasmas movem-se para
as pontas dos cromossomos, ocorrendo a
terminalização dos quiasmas. Figura 7 – Prófase I
21.
20
Metáfase I -As tétrades se distribuem no equador da célula. For-
mando a placa metafásica dupla. A distribuição
em pares nesta etapa permite que ao ocorrer
o encurtamento das fibras cinetocóricas
as mesmas puxem o cromossomo comple-
to (com duas cromátides) para um polo,
não ocorrendo com isso o rompimento
do centrômero e consequentemente a
redução para células haploides ao final da
meiose I.
Anáfase I – O encurtamento das fibras cinetocóricas puxa os
cromossomos homólogos (que estão pareados) para polos opostos
enquanto as fibras polares crescem e distanciam as massas celulares.
Telófase I – Os cromossomos chegam aos polos do fuso e o cito-
plasma se divide.
Fonte das imagens: http://www.infoescola.com
Anáfase I – O encurtamento das fibras cinetocóricas puxa os
cromossomos homólogos (que estão pareados) para polos opostos enquanto
as fibras polares crescem e distanciam as massas celulares.
Telófase I – Os cromossomos chegam aos polos do fuso e o citoplasma
se divide.
http://www.infoescola.com/wp-content/uploads/2009/11/meiose1.jpg
Existe um período entre a Meiose I e a Meiose II, chamado intercinese.
Meiose II. Após o fim da meiose I as células geradas passam por um breve
Anáfase I – O encurtamento das fibras cinetocóricas puxa os
cromossomos homólogos (que estão pareados) para polos opostos enquanto
as fibras polares crescem e distanciam as massas celulares.
Telófase I – Os cromossomos chegam aos polos do fuso e o citoplasma
se divide.
http://www.infoescola.com/wp-content/uploads/2009/11/meiose1.jpg
Existe um período entre a Meiose I e a Meiose II, chamado intercinese.
ocorrer o encurtamento das fibras cinetocóricas as mesmas p
cromossomo completo (com duas cromátides) para um polo, não o
com isso o rompimento do centrômero e consequentemente a redu
células haploides ao final da meiose I.
Figura 8 – Metáfase I
Figura 9 – Anáfase I
Figura 10 – Telófase I
22.
21
Existe um períodoentre a Meiose I e a Meiose II, chamado
intercinese.
Meiose II. Após o fim da meiose I as células geradas passam
por um breve momento denominado intercinese. Neste período, a
carioteca e o nucléolo se refazem, as fibras do fuso desaparecem
e os cromossomos relaxam. Logo em seguida, entram em meiose
II, observe que não ocorre a intérfase, portanto não acontece à
duplicação do material genético nem das estruturas celulares, esse
fato é importante já que a meiose destina-se a gerar gametas (cé-
lulas haploides). A meiose II é dividida em prófase II, metáfase II,
anáfase II e telófase II e os eventos que ocorrem nessas etapas são
semelhantes aos da mitose.
Controle do Ciclo Celular
As multiplicações celulares são controladas continuamente para
evitar que uma célula entre num processo descontrolado de divisões.
As células são orientadas por mecanismos que coordenam os eventos
de síntese no núcleo e no citoplasma e indicam o início e a conclusão
das etapas do ciclo celular. Moléculas controladoras como as ciclinas,
cinases dependentes de ciclinas (CDK, em inglês), fatores de cresci-
mento, proteínas reguladoras e outras, fazem parte do grande leque
de agentes que influenciam e determinam quais passos as células
devem seguir durante o ciclo celular. Esse controle pode ocorrer em
diferentes estágios, e por várias moléculas, abaixo podemos observar
alguns exemplos.
Substâncias Indutoras
Células próximas ou não geram sinais celulares indicando que
uma determinada célula inicie a divisão. Esses sinais podem ser
decorrentes da participação de várias moléculas, entre elas, so-
matomedina, fatores hemopoéticos e o conjunto de substâncias
denominado fatores de crescimento.
23.
22
Normalmente denomina-se ofator de crescimento de acordo com
o local de produção ou célula que irá atuar, podemos exemplificar os
fatores: fibroblástico (FGF), epidérmico (EGF), derivado das plaquetas
(PDGF), hepatócitos (HGF), dos nervos (NGF) e do endotélio vascular
(VEGF). Suas ações são semelhantes entre si, porém podem atuar em
diferentes etapas celulares. Antagônicos aos fatores de crescimentos,
calonas, são substância que inibem a proliferação celular e permitem
assim controlar quando e quanto às células devem se multiplicar.
Ciclinas G1/Cdk2
Boa parte do controle do ciclo celular é devido a interações entre
proteínas ciclinas e CDK. Ciclinas são moléculas produzidas e degra-
dadas rapidamente em diferentes estágios do ciclo celular. Após as
células tomarem a decisão de se dividir e sair da fase G1 (ação esti-
mulada por fatores liberados por outras células), as ciclinas G1 ativam
as cinases Cdk2 (do inglês, cyclin-dependent protein kinase 2), as
quais iniciam um processo de fosforilações que resulta na indução da
replicação do DNA.
O complexo proteico ciclina G1/Cdk2 é denominado de fator
promotor da fase S, SPF (do inglês, S phase-promoting factor) e
será desfeito após a célula dar continuidade na fase S, evitando que
ocorram várias replicações em um mesmo ciclo celular. Durante a G2
existe um processo importante de checagem das moléculas de DNA
recém-sintetizadas, caso seja observado algum erro que não pode
ser reparado, entrarão em ação moléculas que encaminhará à célula
defeituosa a morte.
Ciclinas M/Cdc2
Após as fases de controle durante a intérfase, as células que não
apresentarem empecilhos poderão prosseguir para a divisão, essa
passagem será desencadeada a partir do complexo MPF, fator promo-
tor de mitose (do inglês, M phase-promoting factor) que é formado
24.
23
pelas Ciclinas Munidas a Cdc2 (do inglês, cell-division cycle), similiar
a intérfase, a Cdc2 ativada pelas ciclinas M, inicia um processo de
fosforilação de moléculas intermediárias que regularam vários even-
tos durante a divisão, como: condensação do DNA, formação do fuso
mitótico, desintegração da carioteca, entre outros.
O complexo MPF começa sua degradação apenas quando todos os
cinetócoros estão ligados as suas fibras que vem dos centrossomos,
garantindo assim que ocorrerá uma segregação correta dos cromos-
somos, caso isso não aconteça, sinais são gerados pelo complexo MPF
e a célula interrompe sua divisão.
Supressores Tumorais
Erros no DNA são os principais responsáveis por desencadear a
ativação dos supressores tumorais, proteínas como a p53, p16, p21 e
pRB.Aproteínacitoplasmáticap53ésintetizadaemrespostaapresença
de danos no DNA, a p53 também serve como um fator de transcrição,
induzindo a produção das p16 e p21 que impedem a ação da Cdk2 e
assim bloqueando a replicação do DNA. A pRB também participa em
vários processos de controle do ciclo celular, como na sua fosforilação
que atua com o fator E2F que monitora a multiplicação celular. A inte-
ração entre os supressores com os fatores de crescimentos e com as
ciclinas determinam a progressão de todo o processo da divisão celular
e possibilita o controle e prevenção da difusão de células defeituosas.
25.
24
EXERCÍCIOS
1 – Considerandoque uma espécie possua nº de cromossomas nas células
somáticas 2n=6, a célula apresentada na figura abaixo evidencia esses
cromossomas em:
a) metáfase mitótica.
b) metáfase I.
c) metáfase II.
d) anáfase mitótica.
e) anáfase II
2 – A figura abaixo é característica da Meiose porque só nesse tipo de divisão
celular acontece:
a) separação dos centríolos.
b) formação do fuso acromático.
c) manutenção da carioteca.
d) pareamento dos cromossomos homólogos.
e) duplicação das cromátides.
3 – Quando uma célula conclui a sua primeira divisão meiótica, resultam:
a) duas células diplóides.
b) quatro células diplóides.
c) quatro células haplóides.
d) duas células haplóides.
e) duas células somáticas.
21
Exercícios
1 - Considerando que uma espécie possua nº de cromossomas nas
células somáticas 2n=6, a célula apresentada na figura abaixo evidencia
esses cromossomas em:
a) metáfase mitótica.
b) metáfase I.
c) metáfase II.
d) anáfase mitótica.
e) anáfase II
2 - A figura abaixo é característica da Meiose porque só nesse tipo de
divisão celular acontece:
a) separação dos centríolos.
b) formação do fuso acromático.
21
Exercícios
1 - Considerando que uma espécie possua nº de cromossomas nas
células somáticas 2n=6, a célula apresentada na figura abaixo evidencia
esses cromossomas em:
a) metáfase mitótica.
b) metáfase I.
c) metáfase II.
d) anáfase mitótica.
e) anáfase II
2 - A figura abaixo é característica da Meiose porque só nesse tipo de
divisão celular acontece:
a) separação dos centríolos.
b) formação do fuso acromático.
26.
25
4 – Relacioneas fases meióticas (coluna I) com os respectivos fenômenos
(coluna II):
Coluna I Coluna II
1) zigóteno ( ) Migração dos cromossomos homólogos para os polos
2) paquíteno ( ) pareamento dos homólogos
3) diplóteno ( ) migração dos cromossomos irmãos para os polos.
4) anáfase I ( ) visualização dos quiasmas.
5) anáfase II ( ) ocorrência do crossing-over
A sequência correta, de cima para baixo, na coluna II é
a) 4, 1, 2, 3, 5
b) 4, 1, 5, 2, 3
c) 4, 1, 5, 3, 2
d) 4, 1, 3, 2, 5
e) 4, 2, 5, 1, 3
5 – Uma evidente diferença existente entre a anáfase da mitose e as aná-
fases I e II da meiose é que os cromossomos em migração para os pólos
celulares são:
a) irmãos nas anáfases I e II e homólogos na anáfase da mitose.
b) homólogos nas anáfases I e II e irmãos na anáfase da mitose.
c) homólogos na anáfase I e irmãos na anáfase II e na anáfase da mitose.
d) irmãos na anáfase I e anáfase da mitose e homólogos na anáfase II.
e) irmãos nas anáfases I e II e anáfase da mitose.
27.
26
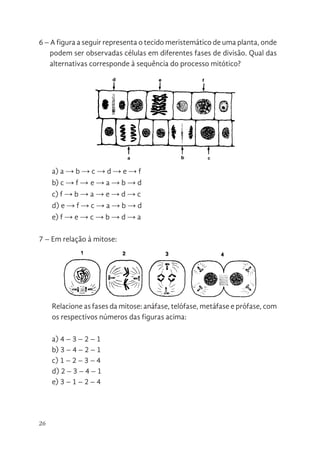
6 – Afigura a seguir representa o tecido meristemático de uma planta, onde
podem ser observadas células em diferentes fases de divisão. Qual das
alternativas corresponde à sequência do processo mitótico?
a) a → b → c → d → e → f
b) c → f → e → a → b → d
c) f → b → a → e → d → c
d) e → f → c → a → b → d
e) f → e → c → b → d → a
7 – Em relação à mitose:
Relacione as fases da mitose: anáfase, telófase, metáfase e prófase, com
os respectivos números das figuras acima:
a) 4 – 3 – 2 – 1
b) 3 – 4 – 2 – 1
c) 1 – 2 – 3 – 4
d) 2 – 3 – 4 – 1
e) 3 – 1 – 2 – 4
23
d) irmãos na anáfase I e anáfase da mitose e homólogos na anáfase II.
e) irmãos nas anáfases I e II e anáfase da mitose.
6 - A figura a seguir representa o tecido meristemático de uma planta,
onde podem ser observadas células em diferentes fases de divisão. Qual
das alternativas corresponde à sequência do processo mitótico?
a) a b c d e f
b) c f e a b d
c) f b a e d c
d) e f c a b d
e) f e c b d a
7 - Em relação à mitose:
Relacione as fases da mitose: anáfase, telófase, metáfase e prófase, com os
respectivos números das figuras acima:
a) 4 - 3 - 2 – 1
b) 3 - 4 - 2 – 1
c) 1 - 2 - 3 - 4
d) 2 - 3 - 4 – 1
e) 3 - 1 - 2 - 4
28.
27
8 – Aconsequência mais importante da mitose é:
a) determinar a diferenciação celular.
b) a produção de gametas e esporos haplóides.
c) a produção de células iguais à célula mãe.
d) aumentar a variabilidade genética dos seres vivos.
e) aumentar a taxa de mutação.
9 – Os ítens abaixo se referem à mitose e todos eles estão corretos, exceto:
a) É um processo de divisão celular importante para o crescimento dos
organismos.
b) Ocorre nas células somáticas de animais e vegetais.
c) Uma célula-mãe origina duas células-filhas com o mesmo número de
cromossomos.
d) A duplicação do DNA ocorre na fase da metáfase.
e) Na fase da telófase, forma-se uma nova membrana nuclear em torno
dos cromossomos e o citoplasma se divide.
10 – Considerando uma célula com 6 cromossomas (2n=6) que esteja em
divisão, o esquema ao lado representaria uma:
a) anáfase I da meiose.
b) metáfase I da meiose.
c) metáfase II da meiose.
d) anáfase II da meiose.
e) anáfase mitótica.
9 - Os ítens abaixo se referem à mitose e todos eles estão corretos,
exceto:
a) É um processo de divisão celular importante para o crescimento dos
organismos.
b) Ocorre nas células somáticas de animais e vegetais.
c) Uma célula-mãe origina duas células-filhas com o mesmo número de
cromossomos.
d) A duplicação do DNA ocorre na fase da metáfase.
e) Na fase da telófase, forma-se uma nova membrana nuclear em torno dos
cromossomos e o citoplasma se divide.
10 - Considerando uma célula com 6 cromossomas (2n=6) que esteja em
divisão, o esquema ao lado representaria uma:
a) anáfase I da meiose.
b) metáfase I da meiose.
c) metáfase II da meiose.
d) anáfase II da meiose.
e) anáfase mitótica.
29.
28
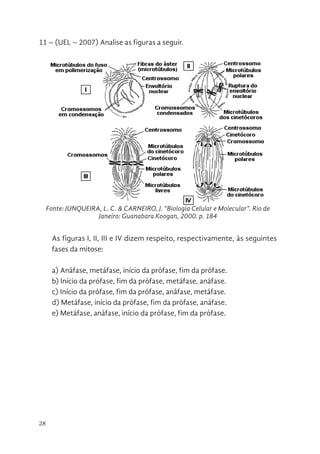
11 – (UEL– 2007) Analise as figuras a seguir.
As figuras I, II, III e IV dizem respeito, respectivamente, às seguintes
fases da mitose:
a) Anáfase, metáfase, início da prófase, fim da prófase.
b) Início da prófase, fim da prófase, metáfase, anáfase.
c) Início da prófase, fim da prófase, anáfase, metáfase.
d) Metáfase, início da prófase, fim da prófase, anáfase.
e) Metáfase, anáfase, início da prófase, fim da prófase.
11 - (UEL – 2007) Analise as figuras a seguir.
Fonte: JUNQUEIRA, L. C. & CARNEIRO, J. "Biologia Celular e Molecular".
Rio de Janeiro: Guanabara Koogan, 2000. p. 184
As figuras I, II, III e IV dizem respeito, respectivamente, às seguintes fases da
mitose:
a) Anáfase, metáfase, início da prófase, fim da prófase.
b) Início da prófase, fim da prófase, metáfase, anáfase.
c) Início da prófase, fim da prófase, anáfase, metáfase.
d) Metáfase, início da prófase, fim da prófase, anáfase.
e) Metáfase, anáfase, início da prófase, fim da prófase.
Fonte: JUNQUEIRA, L. C. & CARNEIRO, J. “Biologia Celular e Molecular”. Rio de
Janeiro: Guanabara Koogan, 2000. p. 184
30.
29
12 – (UFSC– 2005) A Mitose e a Meiose são importantes processos bio-
lógicos, pois permitem que o número de cromossomos de uma célula
permaneça igual, ou seja reduzido, para possibilitar sua restauração
numérica após a fecundação. Com relação aos eventos e aos resultados
destes dois processos, é CORRETO afirmar que:
(01) ao contrário da Mitose, que ocorre em todas as células, a Meio-
se restringe-se àquelas da linha germinativa, que produzirão
gametas.
(02) nos dois processos, ocorre a compactação da cromatina, fenômeno
este que, além de facilitar a divisão correta dos cromossomos, im-
pede que o material genético seja atacado por enzimas, presentes
no citoplasma, que destroem o DNA.
(04) uma mutação que ocorra em uma das cromátides de uma célula
somática será transmitida a todas as suas células-filhas, através
da divisão mitótica.
(08) a Mitose é o sistema de reprodução dos organismos nos quais não
existe a presença de sexo nem a formação de células germinativas.
(16) se considerarmos, em uma mesma espécie, duas células-filhas,
uma originada por Mitose e a outra por Meiose, a primeira conterá
metade do número de cromossomos e o dobro da quantidade de
DNA da segunda.
(32) na Meiose, existe a possibilidade de ocorrer o fenômeno de re-
combinação, que é a troca de segmentos entre quaisquer dois
cromossomos, gerando, com isso, alta variabilidade genética para
os indivíduos envolvidos.
(64) a Meiose compreende duas etapas de divisão cromossômica, sendo
que, após a primeira, o número de cromossomos das células-filhas
é metade do das células-mães.
Soma das alternativas corretas ( )
31.
30
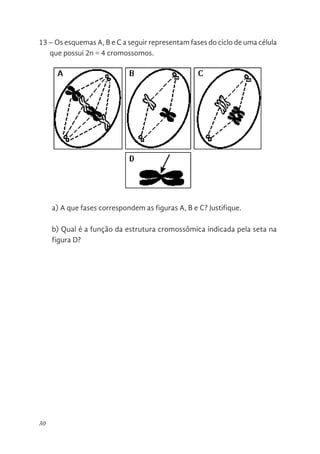
13 – Osesquemas A, B e C a seguir representam fases do ciclo de uma célula
que possui 2n = 4 cromossomos.
a) A que fases correspondem as figuras A, B e C? Justifique.
b) Qual é a função da estrutura cromossômica indicada pela seta na
figura D?
Soma das alternativas corretas ( )
13 - Os esquemas A, B e C a seguir representam fases do c
célula que possui 2n = 4 cromossomos.
a) A que fases correspondem as figuras A, B e C? Justifique.
b) Qual é a função da estrutura cromossômica indicada pela seta n
32.
31
CAPÍTULO II
PRINCÍOS BÁSICOSDE HEREDITARIEDADE
Martín Alejandro Montes e Ana Cristina Lauer Garcia
Nestecapítulofalaremossobreahereditariedadedascaracterísticas
dos seres vivos. Quem primeiro examinou e chegou a conclusões que
geraram os conhecimentos que usamos até hoje foi Gregor Mendel.
O que é genética?
Alguém já comentou que você tem o nariz de seu pai, ou que seus
olhos são iguais aos de sua mãe? Ou ainda, que você se parece com
seus irmãos? Porque será que somos tão parecidos com nossos fami-
liares? Para encontrar as respostas para estas questões necessitamos
recorrer à genética. Esta é a área da biologia que estuda o DNA, nosso
material hereditário encontrado no núcleo de nossas células e em al-
gumas organelas, tais como mitocôndrias e cloroplastos. A Genética
estuda como este material hereditário se transmite de geração em
geração, o modo como ele se expressa e como pode ser alterado ao
longo do tempo.
Gregor Mendel foi o primeiro cientista a descobrir os mecanismos
básicos da hereditariedade e é considerado o “pai” da Genética atual.
Ideias sobre herança antes de Mendel
As questões de como as características se transmitem de geração
a geração sempre despertou curiosidade e muitas hipóteses foram lan-
çadas até a comprovação de suas bases por Gregor Mendel. Uma das
primeiras ideias foi postulada por Anáxagoras de Clazomene (500-428
a.C), o qual defendia que o sêmen produzido pelo homem continha
o protótipo de cada órgão do futuro organismo. Para Anaxágoras, o
papel da mulher seria apenas o de receber e de nutrir o bebê até seu
nascimento. Outra ideia sobre hereditariedade foi lançada pelo filósofo
grego Hipócrates de Cos (460-370 a.C), o qual propôs a hipótese da
pangênese. De acordo com Hipócrates, cada órgão ou parte do corpo
33.
32
de um indivíduoproduziria “gêmulas”, ou partículas hereditárias, que
migrariam para o sêmen do macho e da fêmea. No momento da con-
cepção essas gêmulas se misturariam, formado um novo indivíduo.
Isso explicaria a grande semelhança entre pais e filhos. Outro filósofo
grego, Aristóteles (284-322 a.C), explicava que na formação de um
novo ser o papel da mãe seria o de fornecer a matéria responsável por
nutrir o ser em formação, já o pai, por meio do sêmen, contribuiria
com a “essência”, transmitindo-lhe a alma, fonte da forma e do movi-
mento. Desde a época de Aristóteles muito tempo se passou até que
novas ideias sobre herança fossem estabelecidas. Dando sequência ao
conhecimento de como as características se transmitem de geração
a geração, o médico inglês Willian Harvey (1578-1657) propôs que
todo ser formado dependia da fecundação de um ovo, produzido pela
fêmea, pelo esperma do macho. A partir desta concepção um grupo
de pesquisadores acreditava que neste ovo já existiria um indivíduo
pré-formado, outros acreditavam que este ser estaria abrigado no
espermatozoide.
Quem foi Mendel
Os princípios básicos sobre a heredi-
tariedade,comoconhecidosnosdiasde
hoje, foram estabelecidos por Gregor
Mendel (Figura 1). Nascido em julho
de 1822 na região da Morávia, na atual
República Tcheca, Mendel era filho
de horticultores pobres, o que não
impediu que alcançasse um alto grau
de escolaridade. Em 1843, ingressou
como noviço no monastério agos-
tiniano em Brno. Após a conclusão
de seus estudos, em agosto de 1847,
Mendel foi ordenado padre aos 25 anos
e começou a atuar como professor. Sempre
ão essas gêmulas se misturariam, formado um novo indivíduo. Isso
a a grande semelhança entre pais e filhos. Outro filósofo grego,
es (284-322 a.C), explicava que na formação de um novo ser o papel
seria o de fornecer a matéria responsável por nutrir o ser em formação,
por meio do sêmen, contribuiria com a “essência”, transmitindo-lhe a
nte da forma e do movimento. Desde a época de Aristóteles muito
e passou até que novas ideias sobre herança fossem estabelecidas.
equência ao conhecimento de como as características se transmitem
ção a geração, o médico inglês Willian Harvey (1578-1657) propôs que
formado dependia da fecundação de um ovo, produzido pela fêmea,
erma do macho. A partir desta concepção um grupo de pesquisadores
va que neste ovo já existiria um indivíduo pré-formado, outros
vam que este ser estaria abrigado no espermatozoide.
FOI MENDEL
s princípios básicos sobre a
riedade, como conhecidos nos dias de
am estabelecidos por Gregor Mendel
1). Nascido em julho de 1822 na região
via, na atual República Tcheca, Mendel
o de horticultores pobres, o que não
que alcançasse um alto grau de
dade. Em 1843, ingressou como noviço
astério agostiniano em Brno. Após a
ão de seus estudos, em agosto de
endel foi ordenado padre aos 25 anos
çou a atuar como professor. Sempre
Figura 1. Gregor Mendel
Fonte: Wikipedia (2015)
https://en.wikipedia.org/wiki/File:Gregor_
Mendel_oval.jpg
Figura 1 – Gregor Mendel
Fonte: Wikipedia (2015)
34.
33
destacado em suasfunções, Mendel deu continuidade aos seus estu-
dos na Universidade de Viena, onde aprendeu, entre outras coisas,
a desenhar e executar um trabalho científico. Em 1853, já formado,
retornou para Brno onde desenvolveu, entre 1856 e 1864, suas hoje
famosas experiências com plantas. Em fevereiro de 1865, aos 43 anos,
apresentou os resultados de suas experiências em palestras para a
Sociedade de Ciências Naturais de Brno. O conteúdo apresentado
nestas palestras foi publicado em 1866. Apesar da importância dos
seus experimentos, da qualificação da plateia que assistiu às palestras
e ao interesse geral pelos mecanismos de herança, na época o efeito
dos resultados da pesquisa de Mendel foi mínimo. Ele morreu aos 61
anos, como abade de seu monastério e sem o reconhecimento por
sua notável contribuição à Genética.
A primeira Lei de Mendel
Durante seus estudos Mendel conheceu grandes questões a se-
rem respondidas pela Biologia, entre as quais se destacava a heredi-
tariedade. Visto que até então ninguém havia chegado a uma base
conclusiva sobre o assunto, ele verificou que uma das maneiras de
estudar a hereditariedade seria através de cruzamentos entre varie-
dades que diferissem quanto a características hereditárias com traços
contrastantes. Esses experimentos levaram Mendel a descobrir que as
características são herdadas segundo regras bem definidas e propôs
uma explicação para a existência dessas regras. Seu trabalho genial
colocou-o no nível dos maiores cientistas da humanidade.
Um dos grandes méritos do trabalho de Mendel foi o cuidadoso
planejamento experimental, incluindo a formulação de hipóteses,
análises matemáticas para comprovação de seus resultados e ela-
boração de um mecanismo para explicá-los.
O primeiro cuidado que Mendel teve na realização de seus
experimentos foi com a escolha do material de estudo. As prin-
cipais razões que o levaram a optar pelas ervilhas (Pisum sa-
tivum) foram:
35.
34
1) a facilidadede cultivo;
2) a presença de plantas de caracteres nitidamente distintos e facil-
mente diferenciáveis;
3) a obtenção de descendência fértil a partir do cruzamento entre
variedades distintas;
4) o ciclo de vida curto, o que permite obter várias gerações em pouco
tempo e
5) a facilidade de protegê-las contra polinização estranha, viabilizando
os experimentos com polinização artificial.
As ervilhas são plantas ditas hermafroditas, já que numa mesma
flor há órgãos reprodutores masculinos e femininos. Como estas es-
truturas ficam encobertas, os óvulos de uma flor são fecundados, na
maioria dos casos, por seus próprios grãos de pólen, ocorrendo o que
chamamos de autofecundação.
Inicialmente Mendel selecionou 34 variedades de ervilhas, mas
apenas sete foram selecionadas para seus cruzamentos, já que apre-
sentavam duas formas contrastantes e produção de híbridos férteis
quando cruzadas entre si. Abaixo estão listadas as sete características
estudadas por Mendel.
Características Traços contrastantes
1 Forma da semente Lisa Rugosa
2 Cor da semente (endosperma) Amarela Verde
3 Cor do revestimento da semente Cinza Branca
4 Cor da vagem Amarela Verde
5 Forma da vagem Inflada Estreitada
6 Posição das flores Axial Terminal
7 Altura da planta Alta Anã
Antes de iniciar cada cruzamento, Mendel certificava-se de estar
lidando com plantas de linhagens puras. Podemos definir este termo
como linhagens que geram progênie igual a si ao longo das gerações.
Para Mendel realizar cruzamentos com plantas que naturalmente
36.
35
se autofecundavam, elevaleu-se da técnica de polinização artificial,
realizando o que chamamos de fecundação cruzada. Nesta técnica,
antes do pólen estar maduro e do estigma estar pronto para receber
este pólen, Mendel abria a flor e cortava os estames contendo os
grãos de pólen. Logo após, Mendel fecundava o estigma desta flor
com os grãos de pólen trazidos de outra planta que apresentasse a
característica com o traço desejado.
Mendel tomou o cuidado de analisar apenas uma característica
de cada vez (monohibridismo). Ao cruzar plantas altas com plantas
baixas, por exemplo, ele desconsiderava características como a cor
da semente, a posição das flores no caule, etc.
Mendel iniciou seus experimentos de monohibridismo realizando
a fecundação cruzada de linhagens puras com traços contrastantes,
por exemplo, cruzando uma linhagem pura de sementes lisas com
uma linhagem pura de sementes rugosas. Essa geração é chamada
de “parental” e representada pela letra “P”. Quando os descendentes
da geração parental nasciam formavam a primeira geração de filhos,
ou “F1”. Os descendentes da F1 eram cultivados e autofecundados,
dando origem a segunda geração de filhos, ou “F2”.
Em relação aos resultados dos cruzamentos, Mendel observou que
os indivíduos que formavam a F1 apresentavam somente um dos traços
de uma linhagem parental, ou seja, os indivíduos da F1 eram todos
iguais a mãe ou ao pai. Por exemplo, o cruzamento de plantas puras
de sementes lisas com plantas puras de sementes rugosas sempre
resultava em plantas de sementes lisas. Esse resultado era observado
tanto quando o pólen da geração parental era de plantas de sementes
lisas e o óvulo de plantas de sementes rugosas ou vice-versa. Em vista
destes resultados Mendel se perguntou: será que um dos traços (neste
caso, sementes rugosas) havia desaparecido?
Para responder a este questionamento, Mendel cultivou as se-
mentes da F1 e permitiu que as plantas produzidas sofressem autofe-
cundação, formando a F2. Ao contabilizar a F2 Mendel obteve dois
grandes resultados. O primeirofoi que as sementes rugosas voltaram
37.
36
a aparecer eeste padrão se mantinha nos cruzamentos envolvendo
todas as características analisadas. Assim, segundo Mendel, havia
um traço que ficava em recesso na geração F1 e reaparecia na
geração F2. Com base neste resultado, Mendel denominou o traço
que desaparece na F1 de recessivo e o traço que não desaparece
na F1 de dominante.
Essa observação permitiu que Mendel avançasse em suas conclu-
sões, o traço recessivo não desaparece nem se dilui na F1, simplesmente
o que ocorre é que não se manifesta, porém a informação deste traço
se mantém presente. A grande conclusão deste fato é que, nas palavras
de Mendel, existem “fatores” para cada traço de cada característica
e esses fatores são puros e indissolúveis. Assim surge a ideia de que a
herança é particulada.
O segundo grande resultado de Mendel foi que a proporção de
indivíduos manifestando o traço dominante na F2 era sempre de apro-
ximadamente ¾ (75%) e a proporção de indivíduos manifestando o
traço recessivo era sempre de ¼ (25%). Simplificando, essa relação
era sempre de aproximadamente três indivíduos manifestando o
traço dominante para um indivíduo manifestando o traço recessivo,
ou simplesmente 3:1.
UmresumodetodososresultadosobtidosporMendel,analisandoas
sete características estudadas em ervilhas, pode ser visualizado abaixo.
Característica estudada e
cruzamento de linhagens pu-
ras da geração parental (P) Resultado da F1
Resultado da F2
(autofecunda-
ção da F1)
Propor-
ção
da F2
Forma da semente
Lisa X rugosa
100% lisas
lisas
1.850 rugosas
2,96:1
Cor da semente
Amarela X verde
100% amarelas
6.022 amarelas
2.001 verdes
3,01:1
Cor do revestimento
da semente
Cinza X branca
100% cinzas
705 cinzas
224 brancas
3,15:1
38.
37
Característica estudada e
cruzamentode linhagens pu-
ras da geração parental (P) Resultado da F1
Resultado da F2
(autofecunda-
ção da F1)
Propor-
ção
da F2
Cor da vagem
Verde X amarela
100%verdes
428 verdes
152 amarelas
2,82:1
Forma da vagem
Inflada x estreitada
100% inflada
882 infladas
299 estreitadas
2,95:1
Posição das flores
Axial X terminal
100% axial
651 axiais
207 terminais
3,14:1
Altura da planta
Alta X anã
100% altas
787 altas
277 anãs
2,84:1
Essa constância nos resultados observados nos diferentes cru-
zamentos levou Mendel a pensar na existência de uma lei geral,
responsável pelo mecanismo de herança das ervilhas. Com base em
seus resultados foi elaborada a Primeira Lei de Mendel que postula
o seguinte:
1) cada característica hereditária é determinada por um par de fatores,
transmitidos em igual quantidade pelo pai e pela mãe.
2) Os fatores de cada par separam-se quando os indivíduos produzem
os gametas.
De acordo com estes princípios podemos interpretar os cruzamen-
tos realizados por Mendel. Por exemplo, quando uma planta pura de
semente lisa é cruzada com uma planta pura de semente rugosa, a
primeira apresentará dois fatores para o traço liso e a segunda planta
dois fatores para o traço rugoso. Na produção de gametas a planta
de semente lisa passará para o filho apenas um fator do traço liso e a
rugosa transmitirá apenas um fator o traço rugoso. Desta forma, os
indivíduos híbridos da geração F1 serão todos lisos, mas possuirão um
fator para o traço liso e um fator para o traço rugoso. Esse indivíduo
poderá formar dois tipos de gametas na mesma proporção: ou seja,
39.
38
metade dos gametasconterá um fator para o traço liso e a outra
metade um fator para o traço rugoso. A autofecundação da F1
formará uma F2 contendo uma proporção de três plantas lisas
para uma rugosa (Figura 2).
Os fatores mendelianos são representados nos cruzamentos
por letras, usando a forma maiúscula para o fator que determina
o fenótipo dominante e a minúscula para o fator que determina
o fenótipo recessivo.
Indivíduos que apresentam dois fatores iguais para uma dada
característica são ditos homozigotos, tratam-se dos indivíduos
RR ou rr ilustrados na Figura 2. Indivíduos que possuem dois fa-
tores diferentes para uma característica, por exemplo, os Rr, são
ditos heterozigotos. Ao contrário dos indivíduos homozigotos, os
35
metade dos gametas conterá um fator para o traço liso e a outra metade um
ator para o traço rugoso. A autofecundação da F1 formará uma F2 contendo
ma proporção de três plantas lisas para uma rugosa (Figura 2).
Os fatores mendelianos são representados nos cruzamentos por letras,
sando a forma maiúscula para o fator que determina o fenótipo dominante e a
minúscula para o fator que determina o fenótipo recessivo.
Indivíduos que apresentam dois fatores iguais para uma dada
aracterística são ditos homozigotos, tratam-se dos indivíduos RR ou rr
ustrados na Figura 2. Indivíduos que possuem dois fatores diferentes para
Figura 2. Esquema de um cruzamento monohíbrido de Mendel,
considerando a característica forma da semente de ervilha.
Figura 2. Esquema de um cruzamento monohíbrido de Mendel, considerando a
característica forma da semente de ervilha.
40.
39
heterozigotos podem formardois tipos de gametas, neste caso,
gametas R ou gametas r.
A primeira Lei de Mendel hoje
Na época de Mendel pouco se sabia sobre a transmissão das ca-
racterísticas de geração para geração e houve muitas mudanças nas
terminologias utilizadas em seus trabalhos até a atualidade. O termo
gene, por exemplo, não era conhecido por Mendel. Sabemos que nos
cruzamentos para uma dada característica (monohíbridos), Mendel
estava trabalhando com características condicionadas por um único
gene. Desta forma, podemos definir gene como um fator herdado
que condiciona uma característica. É valido destacar aqui que muitas
características (a grande maioria) são condicionadas por mais de um
gene, é o que chamamos de herança poligênica. Num sentido mais
amplo, gene poderia ser definido como um fragmento de material
genético que codifica para uma função.
Hoje também sabemos que um gene pode existir em diferentes
estados, os alelos. Mendel desconhecia este termo e se referia aos
alelos como fatores (ou traços). Assim, o gene que codifica para a
característica textura da semente da ervilha, ocorre em dois estados,
ou seja, apresenta dois alelos, liso (R) e rugoso (r).
Os alelos são encontrados em um local específico do cromossomo,
chamadodelócus.Alelosdiferentesparaumdeterminadogeneocupam
o mesmo lócus em cromossomos homólogos. Cromossomos homó-
logos são aqueles que apresentam alelos lado a lado, codificando para
as mesmas características. Indivíduos com dois alelos iguais para um
determinado lócus são ditos homozigotos e com dois alelos diferentes
para um dado lócus são denominados de heterozigotos.
Na terminologia atual também podemos definir o termo genótipo
como a soma de todos os alelos de todos os genes de um organismo.
Estamosnosreferindoaogenótipoquandomencionamos,porexemplo,
que uma planta de ervilha é homozigota dominante (RR) ou heterozi-
gota (Rr) em relação à forma da semente. O termo fenótipo pode ser
41.
40
compreendido como amanifestação do genótipo, através do produto
da expressão gênica em um dado ambiente, neste caso, o fenótipo das
ervilhas quanto à forma da semente seria “Lisa” ou “Rugosa”.
Hoje podemos compreender os resultados de Mendel quando ana-
lisamos o comportamento dos cromossomos e dos alelos na meiose,
a divisão celular que leva a formação dos gametas. Esta interpretação
pode ser visualizada na Figura 3. Nesta Figura é possível observar os
alelos responsáveis pela determinação da forma da semente da ervi-
lha, representados pelas letras R e r. Na primeira divisão da meiose,
na anáfase I, os cromossomos homólogos se separam e, consequen-
temente, os alelos de um determinado gene também. Na segunda
divisão da meiose, durante a anáfase II, são as cromátides-irmãs que
se separam. Assim, um indivíduo Rr produz metade dos gametas com
o alelo R e a outra metade com o alelo r.
do genótipo, através do produto da expressão gênica em um dado ambiente,
neste caso, o fenótipo das ervilhas quanto à forma da semente seria “Lisa” ou
“Rugosa”.
Hoje podemos compreender os resultados de Mendel quando
analisamos o comportamento dos cromossomos e dos alelos na meiose, a
divisão celular que leva a formação dos gametas. Esta interpretação pode ser
visualizada na Figura 3. Nesta Figura é possível observar os alelos
responsáveis pela determinação da forma da semente da ervilha,
representados pelas letras R e r. Na primeira divisão da meiose, na anáfase I,
os cromossomos homólogos se separam e, consequentemente, os alelos de
um determinado gene também. Na segunda divisão da meiose, durante a
anáfase II, são as cromátides-irmãs que se separam. Assim, um indivíduo Rr
produz metade dos gametas com o alelo R e a outra metade com o alelo r.
Uma forma fácil de representar e de entender os cruzamentos realizados
Figura 3. Comportamento dos cromossomos e dos
alelos (R e r) responsáveis pela característica forma da
semente da ervilha durante a meiose.
Figura 3. Comportamento dos cromossomos e dos alelos (R e r) responsáveis pela
característica forma da semente da ervilha durante a meiose.
42.
41
Uma forma fácilde representar e de entender os cruza-
mentos realizados por Mendel é mediante a utilização do
quadrado de Punnet, desenvolvido pelo geneticista Reginald
C. Punnet em 1917. O quadrado de Punnet é um diagrama
utilizado para prever os resultados de um determinado cru-
zamento. Com ele é possível visualizar os gametas de cada
genitor e os possíveis genótipos resultantes. Através desta
representação, veja como ficariam os resultados dos cruza-
mentos esquematizados na Figura 2.
Cruzamento entre RR e rr
(parentais)
Cruzamento entre Rr e Rr
(F1)
Gametas produzidos
pelo macho
Gametas produzidos
pelo macho
R R R R
Gametas
produzidos
pela
fêmea
r Rr (Liso) Rr (Liso)
Gametas
produzidos
pela
fêmea
R RR (Liso) Rr (Liso)
r Rr(Liso) Rr (Liso) r Rr (Liso) rr (Rugoso)
Cruzamento teste
Na genética, muitas vezes, é importante saber se um indi-
víduo que apresenta o fenótipo dominante é homozigoto ou
heterozigoto, por exemplo, será que o indivíduo de sementes
lisas é RR ou Rr? Esse questionamento pode ser resolvido pelo
cruzamento teste, que consiste em cruzar o organismo com
fenótipo dominante, para o qual queremos saber o genótipo,
representado como R-, com um organismo de fenótipo reces-
sivo, o qual sempre terá o genótipo rr. Vejamos a seguir os dois
resultados possíveis para este cruzamento e a interpretação
para cada resultado.
43.
42
Cruzamento teste: R-x rr
Resultado
100% das ervilhas apresen-
tam o fenótipo liso para a
forma da semente
50% das ervilhas apresen-
tam o fenótipo liso para a
forma da semente e 50%
apresentam o fenótipo ru-
goso
Interpretação
O indivíduo R- é RR
O indivíduo RR produz
apenas gametas “R” e o in-
divíduo rr produz apenas
gametas com o alelo “r”.
Neste caso a união destes
dois tipos de gametas for-
mará apenas indivíduos Rr
de fenótipo liso para a forma
das sementes.
O indivíduo R- é Rr
O indivíduo Rr formará me-
tade dos gametas com o
alelo “R” e a outra metade
com o alelo “r”. O indivíduo
rr só formará gametas com
o alelo “r”. Logo 50% dos
indivíduos formados será
Rr de fenótipo liso e a outra
metade será rr de fenótipo
rugoso.
Assim, olhando a progênie do cruzamento teste poderemos saber
se o indivíduo dominante é homozigoto ou heterozigoto.
A segunda Lei de Mendel
Mendel estendeu seu trabalho, produzindo cruzamentos entre
linhagens puras para duas características (cruzamentos dihíbri-
dos). Por exemplo, ele estabeleceu uma linhagem pura de forma
de semente lisa (RR) e cor do revestimento da semente cinza (BB)
e outra linhagem pura de forma da semente rugosa (rr) e cor do
revestimento da semente branco (bb). A partir do cruzamento des-
tas linhagens como progenitores (P), Mendel obteve uma F1 onde
44.
43
todos os indivíduosapresentavam sementes lisas e cinzas (RrBb).
A geração F2, obtida pela autofecundação destas plantas da F1,
não apresentava mais a proporção de 3/4 dominantes para 1/4
recessivos. Agora a relação foi de 9/16 para sementes com ambas
as características dominantes (lisas e cinzas), 3/16 para uma carac-
terística dominante e outra recessiva (lisas e brancas), 3/16 para a
outra característica dominante e outra recessiva (rugosas e cinzas)
e 1/16 para as duas características recessivas (rugosas e brancas).
Na Figura 4 estão esquematizados estes resultados.
Podemos visualizar o resultado do cruzamento da F1 (RrBb x RrBb)
em um quadrado de Punnet como a seguir:
Figura 4. Esquema dos cruzamentos dihíbridos de Mendel considerando a
característica forma e cor da semente de ervilha.
e brancas), 3/16 para a outra característica dominante e outra recessiva
(rugosas e cinzas) e 1/16 para as duas características recessivas (rugosas e
brancas). Na Figura 4 estão esquematizados estes resultados.
Podemos visualizar o resultado do cruzamento da F1 (RrBb x RrBb) em
um quadrado de Punnet como a seguir:
Figura 4. Esquema dos cruzamentos dihíbridos de Mendel considerando a
característica forma e cor da semente de ervilha.
45.
44
Gametas produzidos
pelo macho
RBRb rB Rb
Gametas
produzidos
pela
fêmea
RB
RRBB
(lisa/cinza)
RRBb
(lisa/cinza)
RrBB
(lisa/cinza)
RrBb
(lisa/cinza)
Rb
RRBb
(lisa/cinza)
RRbb
(lisa/banca)
RrBb
(lisa/cinza)
Rrbb
(lisa/branca)
rB
RrBB
(liso/cinza)
RrBb
(lisa/cinza)
rrBB
(rugosa/
cinza)
rrBb
(rugosa/
cinza)
rb
RrBb
(lisa/cinza)
Rrbb
(lisa/branca)
rrBb
(rugosa/
cinza)
rrbb
(rugosa/
branca)
Genótipo
(Fenótipo)
Genótipo
(Fenótipo)
Genótipo
(Fenótipo)
Genótipo
(Fenótipo)
Com base nos resultados obtidos no cruzamento ilustrado na
Figura 4 e em outros cruzamentos, Mendel elaborou a hipótese
de que, na formação dos gametas de plantas contrastantes quan-
to a duas características, os fatores de uma dada característica
separam-se independentemente dos fatores que condicionam a
outra característica.
Estas observações permitiram a elaboração da Segunda Lei de
Mendel, ou lei da segregação independente, que afirma que durante
a formação dos gametas, a segregação ou separação dos fatores
de uma característica (hoje chamados de alelos) é independente
da separação dos alelos da outra característica, ou seja, do outro
gene. De acordo com este princípio, a separação dos fatores R e
r ocorrem de forma independente da separação dos outros dois
fatores, B e b. De modo que um indivíduo RrBb forma, em igual
quantidade, os gametas RB, Rb, rB e rb.
46.
45
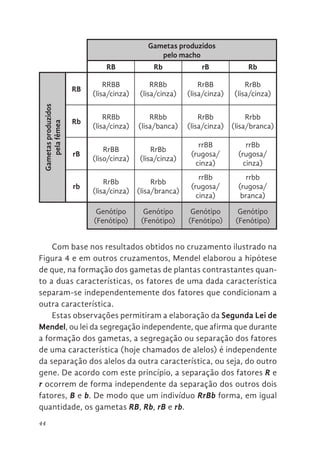
A segunda Leide Mendel Hoje
A Segunda Lei de Mendel se aplica a características codifi-
cadas por genes situados em pares de cromossomos homólogos
diferentes. Cada par de cromossomos homólogos separa-se, in-
dependentemente de todos os outros pares, durante a anáfase I
da meiose. Desta forma, se os alelos do gene que codifica para a
forma da semente de ervilha estiverem localizados em um par de
homólogos diferente do par de homólogos que condiciona a cor
da semente, a segregação destes homólogos ocorrerá de forma
independente e, consequentemente, a separação dos alelos dos
dois genes também ocorrerá de forma independente.
No entanto, se os genes que condicionam a forma e a cor da
semente estiverem em um mesmo par de cromossomos homólo-
gos estes migrarão juntos durante a anáfase I e ficarão no mesmo
gameta, a menos que ocorra crossing-over (ou recombinação)
entre eles. Desta forma, a Segunda Lei de Mendel, não é tão geral
quanto Mendel acreditava, sendo válida somente para casos em
que os genes estão em cromossomos homólogos diferentes ou
quando a localização destes genes num mesmo cromossomo é
distante a ponto de ocorrer crossing-over entre eles.
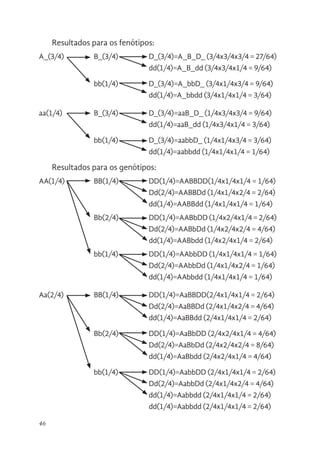
Diagrama ramificado
Quando estamos diante de um cruzamento com mais de dois
genes com dois alelos o quadrado de Punnet fica muito grande
e pode-se utilizar uma forma mais simples para se visualizar os
resultados destes cruzamentos, trata-se do diagrama ramificado.
No esquema abaixo está representado um diagrama ramificado
para três genes com dois alelos e dominância completa nos três
genes (AaBbDd)
Cruzamento: AaBbDd x AaBbDd
47
aa(1/4) BB(1/4) DD(1/4)=aaBBDD(1/4x1/4x1/4= 1/64)
Dd(2/4)=aaBBDd (1/4x1/4x2/4 = 2/64)
dd(1/4)=aaBBdd (1/4x1/4x1/4 = 1/64)
Bb(2/4) DD(1/4)=aaBbDD (1/4x2/4x1/4 = 2/64)
Dd(2/4)=aaBbDd (1/4x2/4x2/4 = 4/64)
dd(1/4)=aaBbdd (1/4x2/4x1/4 = 2/64)
bb(1/4) DD(1/4)=aabbDD (1/4x1/4x1/4 = 1/64)
Dd(2/4)=aabbDd (1/4x1/4x2/4 = 1/64)
dd(1/4)=aabbdd (1/4x1/4x1/4 = 1/64)
O entendimento da genética avançou depois dos trabalhos pio-
neiros de Mendel, seus estudos foram fundamentais para a biologia
e para as ciências como um todo. A organização, o planejamento,
a quantificação e os testes de seus resultados foram pioneiros para
a época, sendo as informações geradas por Mendel a base da ge-
nética atual.
49.
48
EXERCÍCIOS
1 – Comentesobre as razões da ervilha ter sido um organismo tão apropriado
para os estudos de Mendel sobre hereditariedade.
2 – Como era realizada a polinização artificial de ervilhas com traços con-
trastantes?
3 – Defina os seguintes termos:
a) Dominante
b) Recessivo
c) Homozigoto
d) Heterozigoto
e) Genótipo
f) Fenótipo
4 – Na geração parental Mendel cruzava plantas puras de traços contrastan-
tes para uma determinada característica e gerava, na F1, plantas híbridas
com o fenótipo de um dos parentais. Explique o que são plantas puras e
híbridas de acordo com Mendel.
5 – Como Mendel interpretou a proporção de 3:1 obtida na F2 de seus cru-
zamentos com ervilhas?
6 – A finalidade do cruzamento-teste é determinar:
a) o número de genes responsáveis por uma característica monogênica
b) o padrão de herança de uma característica monogênica.
c) a recessividade de uma característica.
d) o grau de penetrância de uma característica.
e) a homozigose ou a heterozigose de um fenótipo dominante.
50.
49
7 – Ocruzamentos de dois indivíduos Aa produzirá descendência consti-
tuída por:
a) 100% de indivíduos AA.
b) 75% de indivíduos AA e 25% aa.
c) 50% de indivíduos AA e 50% aa.
d) 25% de indivíduos AA, 25% aa e 50% Aa.
e) 100% de indivíduos aa.
8 – Os genes A e B estão em cromossomos diferentes. No cruzamento entre
indivíduos duplo heterozigotos AaBb espera-se obter:
a) apenas indivíduos AaBb.
b) indivíduos AB e ab na mesma proporção.
c) indivíduos AABB e aabb, na proporção de 3:1, respectivamente.
d) indivíduos A-B-, A-bb, aaB- e aabb, na proporção de 9:3:3:1, respec-
tivamente
e) indivíduos A-B-, A-bb, aaB- e aabb, na mesma proporção.
9 – Dentre as afirmações abaixo é incorreto afirmar que:
a) Durante a meiose, na formação dos gametas, o número de cromosso-
mos é reduzido à metade e, depois, na fecundação, restabelece-se o
número inicial. Isto garante a constância do número de cromossomos
de uma espécie ao longo das gerações.
b) De acordo com a Primeira Lei de Mendel cada característica é de-
terminada por um par de fatores que se separam na formação dos
gametas, indo um fator do par para cada gameta.
c) Quando os alelos de uma característica são iguais fala-se em condição
heterozigótica (para a qual Mendel usava o termo puro) e, quando os
alelos são diferentes, fala-se em condição homozigótica (para a qual
Mendel usava o termo híbrido).
d) Uma mesma característica pode apresentar duas ou mais formas.
Cada forma é denominada de fenótipo.
e) O termo genótipo pode ser aplicado tanto ao conjunto total de genes
de um indivíduo como a cada gene em particular.
51.
50
10 – Deacordo com a Segunda Lei de Mendel os fatores para duas ou mais
características segregam-se de maneira independente, distribuindo-se
para os gametas ao acaso. De acordo com essa lei, podemos concluir que
um indivíduo de genótipo AABb terá gametas:
a) A, B e b.
b) AA e Bb.
c) AB e Ab.
d) AA, AB, Ab e Bb.
e) Aa e Bb.
52.
51
CAPÍTULO III
VARIAÇÃO ALÉLICAE FUNCIONAMENTO GÊNICO
Maria de Mascena Diniz Maia e Hildson Dornelas Angelo da Silva
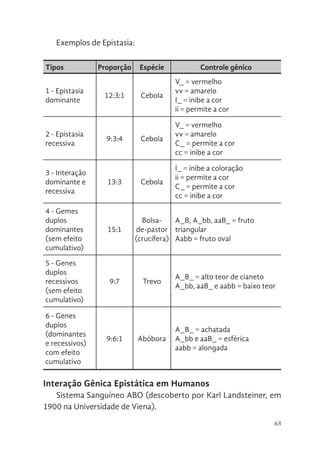
Variação alélica e funcionamento gênico
Os genes podem existir em mais de dois estados alélicos, e cada
alelo pode ter um efeito diferente sobre o fenótipo. Os experimentos
de Mendel estabeleceram que os genes existem em formas alternati-
vas. Para cada uma das sete características que ele estudou – cor da
semente, textura da semente, altura da planta, cor da flor, posição da
flor, forma da vagem e cor da vagem –, Mendel identificou dois alelos,
um dominante e outro recessivo.
Dominância incompleta
Um alelo é dominante se tiver o mesmo efeito fenótipo em he-
terozigotos que em homozigotos, isto é, os genótipos Aa e AA são
fenotipicamente indistinguíveis. Às vezes, entretanto, um heterozi-
goto tem um fenótipo diferente daquele de ambos os homozigotos
associados. A cor da flor maravilha, Mirabilis jalapa, é um exemplo. As
variedades brancas e vermelhas são homozigotas para alelos diferentes
de um gene determinante de cor; quando cruzadas, elas produzem
heterozigotos que tem flores rosa.
O alelo para cor vermelha (W) é, portanto, considerado incom-
pletamente, ou parcialmente dominante em relação ao alelo para a
cor branca (w). A explicação mais provável é que a intensidade de
pigmentação nesta espécie depende da quantidade de um produto
especificado pelo gene para cor. Se o alelo W especifica este produto
e o alelo w não, os homozigotos WW terão o dobro do produto que
os heterozigotos Ww e portanto apresentarão uma cor mais intensa.
Quando o fenótipo do heterozigoto é intermediário aos fenótipos
dos dois homozigotos, como aqui, o alelo parcialmente dominante às
vezes é dito semidominante (da palavra latina pra “metade”,e portanto
semidominante).
53.
52
Fonte: http://bioanatomiaappu.blogspot.com.br/
2013/07/codominancia-y-dominancia-incompleta.html
Fenótipo GenótipoQuantidade do produto gênico
Vermelha WW 2x
Rosa Ww x
Branca ww 0
Codominância
Outra exceção ao princípio de dominância simples surge quando
um heterozigoto apresenta características encontradas em cada um
dos homozigotos associados. Isto ocorre com tipos sanguíneos huma-
nos, que são identificados testando-se produtos celulares especiais
chamados antígenos. Um antígeno é detectado por sua capacidade
de reagir com fatores obtidos do soro do sangue. Esses fatores, que
são produzidos pelo sistema imunológico, reconhecem antígenos
especificamente.
Por exemplo, um soro, anti-M, reconhece apenas o antígeno M nas
células sanguíneas humanas; outro soro anti-N, reconhece apenas o
antígeno N nestas células. Quando um destes soros detecta seu antí-
geno específico em uma tipagem sanguínea, as células aglomeram-se
em uma reação chamada aglutinação. Assim, testando-se células para
Fonte: http://bioanatomiaappu.blogspot.com.br/2013/07/
codominancia-y-dominancia-incompleta.html
Fenótipo Genótipo Quantidade do
produto gênico
Vermelha WW 2x
Rosa Ww x
Branca ww 0
Codominância
Outra exceção ao princípio de dominância simples surge quando um
heterozigoto apresenta características encontradas em cada um do
homozigotos associados. Isto ocorre com tipos sanguíneos humanos, que sã
identificados testando-se produtos celulares especiais chamados antígenos
Um antígeno é detectado por sua capacidade de reagir com fatores obtidos d
soro do sangue. Esses fatores, que são produzidos pelo sistema imunológico
reconhecem antígenos especificamente.
54.
53
aglutinação com sorosdiferentes, pode-se identificar que antígenos
estão presentes e portanto, determinar o tipo sanguíneo.
A capacidade de produzir os antígenos M e N é determinada por
um gene com dois alelos. Um alelo permite que o antígeno M seja
produzido. O outro alelo permite N permite que o antígeno N seja pro-
duzido. Os homozigotos para o alelo M produzem apenas o antígeno
M. Os homozigotos para o alelo N produzem apenas o antígeno N.
Entretanto, os heterozigotos para estes dois alelos produzem ambos
os tipos de antígenos, M e N. Como os dois alelos parecem contribuir
independentemente para o fenótipo dos heterozigotos, eles são cha-
mados codominantes.
A codominância significa que há uma independência de função
dos alelos. Nenhum dos alelos é dominante, ou mesmo parcialmente
dominante, em relação ao outro. Seria, portanto, impróprio distinguir
os alelos por letras maiúsculas e minúsculas, como tivemos em todos
os exemplos anteriores. Em vez disso, os alelos codominantes são
representados por expoentes do símbolo do gene, que neste caso
é a letra L, um tributo a Karl Landsteiner, o descobridor da tipagem
sanguínea. Assim, o alelo M é LM
e o alelo N é LN
.
Genótipo Fenótipo Tipo sanguíneo (antígeno presente)
LM
LM
Antígeno M M
LM
LN
Antígeno MN M N
LN
LN
Antígeno N N
Alelos Múltiplos
O conceito mendeliano de que os genes existem em não mais
que dois estados alélicos teve que ser modificado quando os genes
com três, quatro ou mais alelos foram descobertos. Um exemplo
clássico de um gene com alelos múltiplos é o que controla a cor da
pelagem em coelhos. O gene determinante da cor, indicado pela letra
minúscula c, tem quatro alelos, três dos quais são distinguidos por um
55.
54
expoente: c (albino),ch
(himalaio), cch
(chinchilla) e c+
(tipo selvagem).
Na condição homozigota, cada alelo tem um efeito característico
sobre a cor da pelagem:
cc – pelos brancos por todo o corpo;
ch
ch
– pelos pretos nas extremidades, pelos brancos em todo o
restante;
cch
cch
– pelos brancos com extremidade do corpo pretas;
c+
c+
– pelos coloridos por todo o corpo.
Como a maioria dos coelhos nas populações selvagens são ho-
mozigotos para o alelo c+
,este alelo é chamado tipo selvagem. Em
genética, é costume representar os alelos tipo selvagem por um ex-
poente mais após a letra do gene. Quando o contexto é claro, a letra
às vezes é omitida, e é usado apenas o sinal mais, c+
pode ser abreviado
simplesmente por um +.
Os outros alelos para o gene c são mutantes – formas alteradas
do alelo tipo selvagem que devem ter surgido durante a evolução
do coelho. Os alelos himalaio e chinchilla são indicados por expoen-
tes, mas o alelo albino é indicado simplesmente pela letra c (para
incolor, outro termo para a condição albina). Esta notação reflete
outro hábito na nomenclatura genética: os genes são geralmente
denominados por um alelo mutante, em geral o alelo associado ao
fenótipo mais anormal. A convenção de denominar um gene pelo
alelo mutante é em geral consistente com a convenção. Denominar
os genes pelo alelo recessivo, pois a maioria dos alelos mutantes
são recessivos. Entretanto, às vezes um alelo mutante é dominante,
em cujo caso o gene é dominado por seu fenótipo associado. Por
exemplo, um gene em camundongo controla o tamanho da cauda.
O primeiro alelo mutante deste gene que foi descoberto causa um
encurtamento da cauda em heterozigotos. Este mutante dominante
foi, portanto, simbolizado por T, para tamanho da cauda. Todos os
outros alelos deste gene, e existem muitos, são indicados por uma
56.
55
letra maiúscula eminúscula, dependendo de serem dominantes ou
recessivos; alelos diferentes são distinguidos uns dos outros por
expoentes.
Tipo Genótipo Fenótipo
Albino cc Pelos brancos por todo o corpo
Himalaio ch
ch Pelos pretos nas extremidades, pelos brancos
no restante do corpo
Chinchila cch
cch
Pelos brancos com pontas pretas no corpo
Selvagem c+
c+
Pelos coloridos por todo o corpo
Outro exemplo de alelos múltiplos vem do estudo de tipos san-
guíneos humanos. Os tipos sanguíneos A, B, AB e O, como os tipos
sanguíneos M, N e MN já discutidos, são identificados testando-se
uma amostra de sangue com soros diferentes. Um soro detecta
o antígeno A; outro, o antígeno B. Quando apenas o antígeno A
está presente nas células, o tipo sanguíneo é A; quando apenas
o antígeno B está presente, o sangue é tipo B. Quando ambos os
antígenos estão presentes, o sangue é tipo AB e, quando nenhum
antígeno está presente, o tipo é O. O tipo sanguíneo para os antíge-
nos A e B é totalmente independente dos tipos de antígenos M e N.
O gene responsável por produzir os antígenos A e B é indicado
pela letra I. Este gene tem três alelos, IA
, IB
e i. O alelo IA
especifica
a produção do antígeno A e o alelo IB
especifica a produção do
antígeno B. Entretanto, o alelo i não especifica um antígeno. Entre
os seis genótipos possíveis, existem quatro fenótipos distinguí-
veis – os tipos sangüíneos A, B, AB e O. Neste sistema, os alelos
IA
e IB
são co-dominantes, pois cada um é expresso igualmente
nos heterozigotos IA
IB
, e o alelo i é recessivo tanto em relação
ao alelo IA
quanto ao IB
. Todos os três alelos são encontrados em
frequências apreciáveis em populações humanas; assim, o gene
I é dito polimórfico, para a palavra grega que significa “tendo
muitas formas”.
57.
56
Genótipo Fenótipo AntígenoA Antígeno B Anticorpos no plasma
IAIA ou IAi A + - Anti-B
IBIB ou IBi B - + Anti-B
IA IB AB + + Nenhum
ii O - - Anti-A e Anti-B
Fonte:
http://www.biologiatotal.com.br/blog/falso+o,+efeito+bombaim-150
Por que Algumas Mutações
são Dominantes e Outras Recessivas?
A descoberta de que os genes especificam polipeptídios permitiu
a compreensão da natureza das mutações dominantes e recessivas.
Mutações dominantes têm efeitos fenotípicos em heterozigotos bem
como em homozigotos, enquanto mutações recessivas têm estes efei-
tos apenas em homozigotos. Mutações recessivas em geral envolvem
perda de função do gene, isto é, quando o gene não especifica um
polipeptídio ou quando ele especifica um polipeptídio não funcional
Fonte: http://www.biologiatotal.com.br/blog/falso+o,+efeito+bombaim-150
Por que Algumas Mutações são Dominantes e Outras Recessivas?
A descoberta de que os genes especificam polipeptídios permit
compreensão da natureza das mutações dominantes e recessivas. Mutaç
dominantes têm efeitos fenotípicos em heterozigotos bem como
homozigotos, enquanto mutações recessivas têm estes efeitos apenas
homozigotos. Mutações recessivas em geral envolvem perda de função
IA
IA
ou IA
i A + - Anti-B
IB
IB
ou IB
i B - + Anti-B
IA
IB
AB + + Nenhum
ii O - - Anti-A e Ant
58.
57
ou subfuncional. Asmutações recessivas são, portanto, tipicamente
alelos de perda de função. Tais alelos têm pouco ou nenhum efeito
discernível na condição heterozigota com um alelo tipo selvagem por-
que o alelo selvagem especifica um polipeptídio funcional que fará seu
papel normal no organismo. O fenótipo de um heterozigoto mutante/
selvagem será,portanto, o mesmo, ou essencialmente o mesmo, que
o do homozigoto tipo selvagem.
Algumas mutações recessivas resultam em perda parcial de
função do gene. Por exemplo, o alelo himalaio do gene da cor de
pelagem em mamíferos tais como coelhos e gatos especifica um
polipeptídio que funciona apenas nas partes do corpo onde a tem-
peratura é reduzida. Esta perda parcial de função explica porque
animais homozigotos para o alelo himalaio têm pelos pigmentados
em suas extremidades, mas não no resto do corpo. Nas extremidades,
o polipeptídeo especificado por este alelo é funcional, enquanto no
resto do corpo ele não é. A expressão do alelo himalaio é, portanto,
sensível à temperatura.
Algumas mutações dominantes podem também envolver perda
de função gênica. Se o fenótipo controlado por um gene é sensível
à quantidade do produto gênico, uma mutação de perda de função
pode levar a um fenótipo mutante na condição heterozigota com
um alelo tipo selvagem. Outras mutações dominantes interferem
na função do alelo tipo selvagem especificando polipeptídios que
inibem, antagonizam ou limitam a atividade do polipeptídio tipo
selvagem. Tais mutações às vezes são chamadas de mutações do-
minantes negativas.
Algumas mutações dominantes causam o fenótipo mutante na
condição heterozigota com um alelo tipo selvagem porque elas acen-
tuam a função do produto gênico. A função acentuada pode surgir
porque a mutação especifica um polipeptídio novo ou porque faz com
que o polipeptídio tipo selvagem seja produzido onde ou quando ele
não deveria ser. As mutações dominantes que funcionam deste modo
são chamadas mutações de ganho de função.
59.
58
Influência do Ambientena Expressão do Gene
Ambientes biológicos e físicos são fatores que influenciam no fun-
cionamento do gene. Os fatores no ambiente físico são mais fáceis de
estudar,poisdeterminadosgenótipospodemsercriadosnolaboratório
sob condições controladas permitindo uma avaliação dos efeitos de
temperatura, luz, nutrição e umidade. Como exemplo, consideremos a
mutaçãodeDrosophilaconhecidacomoshibire.Natemperaturanormal
de cultura, 25°C, as moscas shibire são viáveis e férteis, mas são extre-
mamentesensíveisaumchoquesúbitodetemperatura.Outroexemplo
é a expressão do alelo himalaio em coelhos que produz pelagem escu-
ra na extremidade do corpo, nariz, orelhas e pés. Porém, o pigmento
escuro só se desenvolve quando o coelho é criado a 25º C ou menos.
Se o coelho é criado a 30ºC, não se desenvolvem as partes escuras. A
expressão do alelo himalaio é, portanto, dependente da temperatura,
a enzima necessária para a produção do pigmento escuro é inativada a
temperaturas mais altas.
Podemos deduzir que os genes e seus produtos não atuam isola-
damente. Por vezes, fatores ambientais isolados podem produzir um
fenótipo que é o mesmo que o fenótipo produzido por um genótipo.
Essefenótipoéoquechamamosdefenocópia.Portanto,umafenocópia
é uma característica produzida por efeitos ambientais que mimetizam
o fenótipo produzido por um genótipo. Por exemplo, em moscas de
frutas, a mutação autossômica recessiva sem olho produz olhos muito
reduzidos. Porém, o fenótipo sem olho também pode ser produzido
pela exposição das larvas de moscas normais ao metaborato de sódio.
Podemos exemplificar também, como o ambiente físico influencia o
fenótipo em humanos. A fenilcetonúria (PKU) é um distúrbio recessivo
dometabolismodoaminoácidofenilalanina.Criançashomozigotaspara
o alelo mutante acumulam substâncias tóxicas em seus cérebros que
podem prejudicar a capacidade mental afetando o desenvolvimento do
cérebro. Outro exemplo, a surdo-mudez não hereditária, em humanos,
que pode decorrer, por exemplo, de infecções graves que atingem o
sistema nervoso, é uma fenocópia da surdo-mudez de causa genética.
60.
59
Penetrância e Expressividade
Podemosver até agora, nos cruzamentos genéticos considerados,
que cada indivíduo com um determinado genótipo expressa o fenótipo
esperado. Porém para algumas características, tal suposição não ocor-
re, ou seja, o genótipo nem sempre produz o fenótipo esperado, um
fenômeno chamado penetrância incompleta. A penetrância é definida
como a percentagem de indivíduos tendo um genótipo particular que
expressa o fenótipo esperado. Por exemplo, a polidactilia humana, a
condição de ter dedos e artelhos extras, uma característica causada por
um alelo dominante. Ocasionalmente, as pessoas em uma família, cujos
filhos herdaram o alelo, um dos filhos tem dedos ou artelhos normais.
Nesse caso o gene para polidactilia não é totalmente penetrante. Por
exemplo se examinarmos 42 pessoas tendo um alelo para polidactilia
e encontrarmos que 30 deles apresentam a característicafenotípica,
a penetrância seria de 30/42= 0,71 (71%).
A expressividade é definida como o grau no qual uma característica
é expressa. A polidactilia além de penetrância incompleta apresenta
também expressividade variável, algumas pessoas possuem dedos e
artelhos extras que são funcionais e outras possuem apenas pequenas
marcas de peles extras. A penetrância incompleta e a expressividade
variável são devidas aos efeitos de outros genes e a fatores ambien-
tais que podem alterar ou suprimir completamente o efeito de um
determinado gene.
Interações Gênicas
A interação gênica ocorre quando vários pares de genes atuam
num único caráter hereditário. Os genes podem ou não estar locali-
zados em um mesmo cromossomo. As primeiras evidências de que
uma característica pode ser influenciada por mais de um gene foram
obtidas por Bateson e Punnett de experimentos de cruzamentos com
galinhas. O trabalho destes pesquisadores demonstrou que dois genes
que segregam independentemente podem afetar uma mesma carac-
terística. Combinações diferentes de alelos dos dois genes resultaram
61.
60
em fenótipos diferentes,supostamente devido a interações de seus
produtos em nível bioquímico ou celular. A interação gênica pode ser
compreendida como não epitática e epistática.
Interação Gênica não Epistática
Como exemplo de interação gênica não epistática, vamos consi-
derar a forma das cristas em galinhas estudada por Bateson e Punnett
que pode manifestar-se com quatro fenótipos diferentes: ervilha,
rosa, noz e simples. Vários cruzamentos entre essas aves permitiram
concluir que o caráter em questão depende da interação entre dois
pares alelos: R e E. Cada um desses pares apresenta um gene que atua
como dominante (R ou E) em relação ao seu alelo recessivo (r ou e).
Os experimentos demonstraram o seguinte tipo de interação:
crista ervilha: manifesta-se na presença do gene E, desde que não
ocorra o gene R.
crista rosa: manifesta-se na presença do gene R, desde que não ocorra
o gene E.
crista noz: manifesta-se quando ocorrem os gene e R.
crista simples: manifesta-se na ausência dos genes E e R.
Assim, podemos considerar os seguintes genótipos para os quatro
fenótipos existentes:
Genótipo Fenótipo
EErr ou Eerr crista ervilha
eeRR ou eeRr crista rosa
EERR, EERr, EeRR ou EeRr crista noz
eerr crista simples
62.
61
Interação Gênica Epistática
Quandoum par de genes inibe outro par que se localiza em cro-
mossomo não homólogo para que este não manifeste seu caráter
hereditário. O gene que inibe é chamado epistático e o gene inibido
é chamado hipostático.
Características da interação gênica epistática:
a. Ocorre quando dois ou mais genes determinam a produção de
enzimasquecatalisamdiferentesetapasdeumamesmaviabios-
sintética:
SP = substância precursora;
S1
e S2
= produtos intermediários;
PF = produto final;
A e B = alelos que produzem enzimas normais;
a e b = alelos que produzem enzimas que causam bloqueio na
via biossintética.
b. Quando a epistasia envolve a supressão gênica inter-alé-
lica, ou seja, os alelos de um loco gênico encobre a expres-
são de outro alelo pertencente a outro loco gênico (não-a-
lelo). Isto pode ser evidenciado na seguinte via, em que:
hereditário. O gene que inibe é chamado epistático e o gene in
chamado hipostático.
Características da interação gênica epistática:
a) Ocorre quando dois ou mais genes determinam a produção de e
que catalisam diferentes etapas de uma mesma via biossintética:
SP = substância precursora;
S1 e S2 = produtos intermediários;
PF = produto final;
A e B = alelos que produzem enzimas normais;
a e b = alelos que produzem enzimas que causam bloqueio
biossintética.
b) Quando a epistasia envolve a supressão gênica inter-alélica, ou
alelos de um loco gênico encobre a expressão de outro alelo perten
outro loco gênico (não-alelo). Isto pode ser evidenciado na seguinte
que:
Características da interação gênica epistática:
a) Ocorre quando dois ou mais genes determinam a produção de en
que catalisam diferentes etapas de uma mesma via biossintética:
SP = substância precursora;
S1 e S2 = produtos intermediários;
PF = produto final;
A e B = alelos que produzem enzimas normais;
a e b = alelos que produzem enzimas que causam bloqueio n
biossintética.
b) Quando a epistasia envolve a supressão gênica inter-alélica, ou se
alelos de um loco gênico encobre a expressão de outro alelo pertence
outro loco gênico (não-alelo). Isto pode ser evidenciado na seguinte vi
que:
SP = determina ausência de cor; (branco)
S1 = determina a cor amarela;
63.
62
SP = determinaausência de cor; (branco)
S1
= determina a cor amarela;
S2
= determina a cor vermelha;
Desta forma tem-se a seguinte ação gênica:
A_: permite a expressão da cor
aa: inibe a cor (branco)
B-: manifesta a cor vermelha
bb: manifesta a cor amarela
Conclusão: neste caso, aa inibe ou suprime a ação do loco Bb:
aa é epistático sobre Bb. Bb é o gene hipostático.
c. Quando se verifica epistasia entre dois locos gênicos, o número
de fenótipos entre os descendentes será menor que quatro. A
proporção 9:3:3:1 se modifica dando origem a uma combinação
daquela proporção.
Exemplos de interação epistática:
Tipo Forma epistática Proporção esperada
1. Epistasia dominante (A) 12:3:1
2. Epistasia recessiva (a) 9:3:4
3. Genes duplos dominantes
com efeito cumulativo
(A,B) 9:6:1
4. Genes duplo dominantes sem
o efeito cumulativo
(A,B) 15:1
5. Genes duplos recessivos com
efeito cumulativo
(a,b) 9:6:1
6. Genes duplos recessivos sem
o efeito cumulativo
(a,b) 9:7
7. Interação dominante e
recessiva
(A,b) 13:3
64.
63
Exemplos de Epistasia:
TiposProporção Espécie Controle gênico
1 - Epistasia
dominante
12:3:1 Cebola
V_ = vermelho
vv = amarelo
I_ = inibe a cor
ii = permite a cor
2 - Epistasia
recessiva
9:3:4 Cebola
V_ = vermelho
vv = amarelo
C_ = permite a cor
cc = inibe a cor
3 - Interação
dominante e
recessiva
13:3 Cebola
I_ = inibe a coloração
ii = permite a cor
C_ = permite a cor
cc = inibe a cor
4 - Gemes
duplos
dominantes
(sem efeito
cumulativo)
15:1
Bolsa-
de-pastor
(crucífera)
A_B, A_bb, aaB_ = fruto
triangular
Aabb = fruto oval
5 - Genes
duplos
recessivos
(sem efeito
cumulativo)
9:7 Trevo
A_B_ = alto teor de cianeto
A_bb, aaB_ e aabb = baixo teor
6 - Genes
duplos
(dominantes
e recessivos)
com efeito
cumulativo
9:6:1 Abóbora
A_B_ = achatada
A_bb e aaB_ = esférica
aabb = alongada
Interação Gênica Epistática em Humanos
Sistema Sanguíneo ABO (descoberto por Karl Landsteiner, em
1900 na Universidade de Viena).
65.
64
Os grupos sanguíneosABO (adaptado de Thompson, 1981)
Grupo sanguíneo
(fenótipo)
Genótipo
Antígenos nos
eritrócitos
Anticorpos
no plasma
O Ii Nenhum Anti-A e anti-B
A IA
IA
ou IA
i A Anti-B
B IB
IB
ou IB
i B Anti-A
AB IA
IB
A e B Nenhum
Os antígenos A e B têm como substrato outro, designado antígeno
H, e são sintetizados pela ação do gene I, que possui três alternativas
alélicas: IA
, IB
e i. Em relação ao alelo i pensa-se que é silencioso, ou
seja, que não têm nenhum produto ativo e desta forma os eritrócitos
de indivíduos homozigóticos para este alelo transportam o antígeno
H inalterado. Um fenótipo muito particular e raro na população, co-
nhecido como “fenótipo de Bombaim”, refere-se aos indivíduos Oh,
os quais não possuem antígenos A, B e H e têm anticorpos anti-A,
anti-B e anti-H. Tendo em conta o pedigree da figura 1, uma mulher
cujo fenótipo é Oh (assinalado a vermelho), descendente de um casa-
mento consanguíneo e casada com um homem do tipo A deu origem
a uma filha AB.
Figura 1 – Genealogia mostrando o fenótipo Oh (Genótipo hh) no indivíduo II-5.
Fonte: Thompson e Thompson, 1988.
Os antígenos A e B têm como substrato outro, designado antígeno H,
são sintetizados pela ação do gene I, que possui três alternativas alélicas: IA
IB e i. Em relação ao alelo i pensa-se que é silencioso, ou seja, que não tê
nenhum produto ativo e desta forma os eritrócitos de indivíduos homozigótico
para este alelo transportam o antígeno H inalterado. Um fenótipo mui
particular e raro na população, conhecido como “fenótipo de Bombaim”, refere
se aos indivíduos Oh, os quais não possuem antígenos A, B e H e tê
anticorpos anti-A, anti-B e anti-H. Tendo em conta o pedigree da figura 1, um
mulher cujo fenótipo é Oh (assinalado a vermelho), descendente de u
casamento consanguíneo e casada com um homem do tipo A deu origem
uma filha AB.
Figura 1. Genealogia mostrando o fenótipo Oh (Genótipo hh) no indivíduo II-5
Fonte: Thompson e Thompson, 1988.
66.
65
Por estudos dosancestrais chegou-se à conclusão que a mulher
possuía o alelo IB
, mas não conseguia formar o antígeno respectivo. Isto
só pode ser explicado pela existência de outro gene envolvido neste
fenótipo: o gene H, para o qual a mulher era homozigotica recessiva.
Desta forma, sem o antígeno H, os alelos IA
e IB
mesmo que presentes
não podem exprimir o fenótipo visto não terem substrato para atuar.
Figura 2 – Hipotética via biossintética dos antígenos A, B e H.
Os antígenos A, B e H são macromoléculas muito semelhantes
entre si, diferindo apenas nos glicídeos terminais que conferem a espe-
cificidade antigênica. Estes glicídeos são adicionados por transferases
(enzimas) a uma cadeia proteica, supondo-se serem estas enzimas
os produtos dos alelos IA
, IB
e H. Com estes dados é fácil entender as
relações entre os dois genes envolvidos – I e H – no sistema sanguí-
neo ABO através de uma hipotética via biossintética destes antígenos
esquematizada na figura 2. Note-se que os indivíduos com o genótipo
‘hh’ não sintetizam nenhum antígeno pois não possuem a enzima H –
transferase (produto do alelo H), mantendo o precursor inalterado. Da
mesma forma, os indivíduos ‘ii’, não conseguem sintetizar antígenos A
e/ou B devido à ausência das enzimas responsáveis pela sua produção.
Posto isto, e com base neste exemplo clássico de interação
gênica no Homem, é de salientar que numa análise biológica das
Por estudos dos ancestrais chegou-se à conclusão que a mulhe
alelo IB, mas não conseguia formar o antígeno respectivo. Isto s
explicado pela existência de outro gene envolvido neste fenótipo:
para o qual a mulher era homozigotica recessiva. Desta forma, sem
H, os alelos IA e IB mesmo que presentes não podem exprimir o fe
não terem substrato para atuar.
Figura 2. Hipotética via biossintética dos antígenos A, B e H.
Os antígenos A, B e H são macromoléculas muito semelhante
diferindo apenas nos glicídeos terminais que conferem a es
antigênica. Estes glicídeos são adicionados por transferases (enzim
cadeia proteica, supondo-se serem estas enzimas os produtos dos
e H. Com estes dados é fácil entender as relações entre os
envolvidos – I e H – no sistema sanguíneo ABO através de uma h
biossintética destes antígenos esquematizada na figura 2. Note
indivíduos com o genótipo ‘hh’ não sintetizam nenhum antígeno
possuem a enzima H – transferase (produto do alelo H), mantendo
inalterado. Da mesma forma, os indivíduos ‘ii’, não conseguem
67.
66
características hereditárias énecessário aprofundar o estudo das
mesmas para não tirar conclusões precipitadas. Tal como a inte-
ração gênica, também outros fatores podem alterar a frequência
fenotípica esperada na descendência, nomeadamente a penetrância
incompleta, a expressividade variável, letalidade, interação com o
ambiente, entre outros.
68.
67
EXERCÍCIOS
1 – EmMirabilis jalapa, planta conhecida pelo nome vulgar de maravilhosa,
existem três tipos de coloração das pétalas das flores: branca, vermelha
e rosa. Ao serem cruzadas plantas de flores brancas com plantas de flo-
res vermelhas, origina-se uma prole 100% rosa. Do cruzamento dessas
últimas obtêm-se, na descendência, plantas com flores rosa, vermelhas
ou brancas. Em relação às características genéticas da Mirabilis jalapa,
assinale a alternativa correta.
a) Existem dois alelos, envolvidos no padrão de coloração dessas flores,
que exibem uma relação típica do codominância.
b) As flores rosa são heterozigotas.
c) As flores brancas e vermelhas são homozigotas.
d) Do cruzamento entre plantas com flores vermelhas e plantas com
flores rosa, espera-se que 50% da descendência apresentem flores
vermelhas e o restante, flores rosa.
e) Da autopolinização de flores brancas poderão originar-se plantas com
flores vermelhas ou rosa.
2 – (UFRN) A planta “maravilha” – Mirabilis jalapa – apresenta duas varieda-
des para a coloração das flores: a Alba (flores brancas) e a rubra (flores
vermelhas). Cruzando as duas variedades, obtêm-se, em F1, somente
flores róseas. Do cruzamento entre duas plantas heterozigotas, qual
distribuição fenotípica da prole?
3 – Uma caixa de coelhos continha uma fêmea himalaia, um macho albino
e um macho chinchila. A fêmea teve oito descendentes: dois himalaias,
quatro chinchilas e dois albinos. Qual coelho foi o pai e quais eram os
genótipos da fêmea, do macho e de seus descendentes?
4 – Em galinhas da raça minorca, a plumagem pode ser branca, preta e cin-
za-azulado. Do cruzamento de dois indivíduos um branco e outro preto,
encontre os genótipos e fenótipos de F1 e F2.
69.
68
5 – Nogado shorthorn existe um par de alelos condicionador da cor da
pelagem, não apresentando dominância entre os alelos (vv = vermelho;
bb = branco; vb = vermelho e branco). Qual a porcentagem esperada
para os descendentes do cruzamento de vaca de pelagem branca com
um touro que, anteriormente cruzado várias vezes com uma vaca de
pelagem vermelha forneceu 50% de descendentes com pelagem ver-
melha e branca e 50% de pelagem vermelha?
a) 50% vermelha e branca e 50% branca.
b) 50% vermelha e 50% vermelha e branca.
c) 75% vermelha e 25% vermelha e branca.
d) 25% vermelha, 25% branca e 25% vermelha e branca.
e) 100% vermelha
6 – Os grupos sanguíneos do sistema MN são: M,N e MN, determinados por
dois alelos de um gene :M e N. Um homem de sangue M, casa-se com
uma mulher de sangue N, sendo uma caso de co-dominância, encontre
os genótipos e fenótipos de F1 e F2.
7 – Num banco de sangue foram selecionados os seguintes doadores: grupo
AB=15; grupo A=25; grupo B=8; grupo O=20. O primeiro pedido de doação
partiu de um hospital que tinha dois pacientes nas seguintes condições:
Paciente I: possui ambos os tipos de aglutininas no plasma.
Paciente II: possui apenas um tipo de antígeno nas hemácias e aglutinina
b no plasma.
Quantos doadores estavam disponíveis para os pacientes I e II, respec-
tivamente?
8 – (UNIMONTES) Na década de 40, uma ação de paternidade envolvendo o
atordecinemaCharlieChaplinfoianalisada,utilizando-seainformaçãosobre
o tipo sanguíneo do sistema ABO dos envolvidos. Veja os dados a seguir:
70.
69
Chaplin – sanguetipo O
Mãe da criança – sangue tipo A
Criança – sangue tipo B
Embora, na época, o júri tivesse dito que Chaplin era o pai da criança, a
análise genética mostra que ele não o poderia ser. Analise as afirmativas
abaixo e assinale a que melhor justifica o fato de o ator não poder ser
o pai da criança.
a) A criança era seguramente heterozigota.
b) A mãe era seguramente homozigota.
c) Chaplin era homozigoto dominante.
d) Nem Chaplin nem a mãe tinham o alelo B herdado pela criança.
9 – (UNESP) Epistasia é o fenômeno em que um gene (chamado epistá-
tico) inibe a ação de outro que não é seu alelo (chamado hipostático).
Em ratos, o alelo dominante B determina cor de pelo acinzentada,
enquanto o genótipo homozigoto bb define cor preta. Em outro cro-
mossomo, um segundo lócus afeta uma etapa inicial na formação dos
pigmentos dos pelos. O alelo dominante A nesse lócus possibilita o
desenvolvimento normal da cor (como definido pelos genótipos B_
ou bb), mas o genótipo aa bloqueia toda a produção de pigmentos e o
rato torna-se albino. Considerando os descendentes do cruzamento de
dois ratos, ambos com genótipo AaBb, os filhotes de cor preta poderão
apresentar genótipos:
a) Aabb e AAbb.
b) Aabb e aabb.
c) AAbb e aabb.
d) AABB e Aabb.
e) aaBB, AaBB e aabb.
71.
70
10 – Acor dos pelos nas cobaias é condicionada por uma série de alelos
múltiplos com a seguinte escala de dominância: C (preta) > C1 (marrom)
> C2 (creme) > c (albino). Uma fêmea marrom teve três ninhadas, cada
uma com um macho diferente. A tabela a seguir mostra a constituição
de cada ninhada. A partir desses dados, qual seria o genótipo e fenótipo
do macho responsável pela ninhada 2?
Número de Descendentes
Ninhada Pretos Marrons Cremes Albinos
1 5 3 0 2
2 0 4 2 2
3 0 5 0 4
11 – (FUVEST) Em cães labradores, dois genes, cada um com dois alelos
(B/b e E/e), condicionam as três pelagens típicas da raça: preta, marrom
e dourada. A pelagem dourada é condicionada pela presença do alelo
recessivo e em homozigose no genótipo. Os cães portadores de pelo
menos um alelo dominante E serão pretos, se tiverem pelo menos um
alelo dominante B; ou marrons, se forem homozigóticos bb. O cruzamento
de um macho dourado com uma fêmea marrom produziu descendentes
pretos, marrons e dourados. O genótipo do macho é:
a) Ee BB.
b) Ee Bb.
c) ee bb.
d) ee BB.
e) ee Bb.
12 – (UFRS) Na cebola, a presença de um alelo dominante C determina a
produção de bulbo pigmentado; em cebolas cc, a enzima que catalisa a
formação de pigmento não é produzida (cebolas brancas). Outro gene,
herdado de forma independente, apresenta o alelo B, que impede a
72.
71
manifestação de geneC. Homozigotos bb não têm a manifestação da
cor do bulbo impedida. Quais as proporções fenotípicas esperadas do
cruzamento de cebolas homozigotas coloridas com BBcc?
a) 9/16 de cebolas brancas e 7/16 de cebolas coloridas.
b) 12/16 de cebolas brancas e 4/16 de cebolas coloridas.
c) 13/16 de cebolas brancas e 3/16 de cebolas coloridas.
d) 15/16 de cebolas brancas e 1/16 de cebolas coloridas.
e) 16/16 de cebolas brancas.
13 – (MACKENZIE) Em galinhas, a cor da plumagem é determinada por 2
pares de genes. O gene C condiciona plumagem colorida enquanto seu
alelo c determina plumagem branca. O gene I impede a expressão do
gene C, enquanto seu alelo i não interfere nessa expressão. Com esses
dados, conclui-se que se trata de um caso de:
a) epistasia recessiva.
b) herança quantitativa.
c) pleiotropia.
d) codominância.
e) epistasia dominante.
14 – A cor do pelo em coelhos depende de uma serie de alelos múltiplos: C,
cch, ch, e ca, responsáveis, respectivamente, pelas pelagens selvagem
(aguti), chinchila, himalaia e albina. A relação de dominância entre eles é
C > cch > ch >ca. Um criador colocou uma fêmea himalaio e dois machos
chinchilas, de genótipos diferentes, em uma mesma gaiola. Depois de um
certo tempo, nasceram 8 coelhinhos, sendo 4 chinchilas, 2 himalaias e 2
albinos. Com base nesses dados, responda:
a)Qualogenótipodafêmeahimalaioedomachochinchilaqueafertilizou?
b) Qual o resultado fenotípico esperado se a mesma fêmea himalaio
cruzar com outro macho chinchila e nascerem outros 8 coelhinhos?
73.
72
15 – (Vunesp-2005)Considere as seguintes formas de herança:
I. Na planta boca-de-leão, há indivíduos homozigotos, cujo genótipo
(CV
CV
) define cor vermelha nas flores. Indivíduos homozigotos com
genótipos (CB
CB
) apresentam flores brancas. Os heterozigotos resul-
tantes do cruzamento entre essas duas linhagens (CV
CB
)apresentam
flores de cor rosa.
II. Em humanos, indivíduos com genótipos IA
IA
ou IA
i apresentam tipo
sangüíneo A e os com genótipos IB
IB
ou IB
i apresentam tipo sangüíneo
B. Os alelos IA
e IB
são, portanto, dominantes com relação ao alelo i.
Por outro lado, o genótipo IA
IB
determina tipo sanguíneo AB.
III. A calvície é determinada por um alelo autossômico. Homens com
genótipo C1
C1
(homozigotos) ou C1
C2
(heterozigotos) são calvos,
enquanto mulheres C1
C1
são calvas e C1
C2
são normais. Tanto homens
quanto mulheres C2
C2
são normais.
I, II e III são, respectivamente, exemplos de:
a) dominância incompleta, co-dominância e expressão gênica influen-
ciada pelo sexo.
b) dominância incompleta, pleiotropia e penetrância incompleta.
c) co-dominância, epistasia e pleiotropia.
d) epistasia, co-dominância e dominância incompleta.
e) herança poligênica, dominância incompleta e expressão gênica in-
fluenciada pelo sexo.
16 – Em camundongos, a coloração da pelagem é determinada por dois pares
de genes, Aa e Cc, com segregação independente. O gene A determina
coloração aguti e é dominante sobre seu alelo a que condiciona coloração
preta. O gene C determina a produção de pigmentos e é dominante sobre
seu alelo c que inibe a formação de pigmentos dando origem a indivíduos
albinos. Do cruzamento de um camundongo preto com um albino, foram
obtidos apenas descendentes agutis. Qual é o genótipo desse casal?
74.
73
a) aaCC xaacc
b) Aacc x aaCc
c) aaCc x AAcc
d) AaCc x AaCc
e) aaCC x AAcc
17 – (UFU) A cor da pelagem em cavalos depende, dentre outros fatores, da
ação de dois pares de genes Bb e Ww. O gene B determina pelos pretos
e o seu alelo b determina pelos marrons. O gene dominante W “inibe”
a manifestação da cor, fazendo com que o pelo fique branco, enquanto
que o alelo recessivo w permite a manifestação da cor.
Cruzando-se indivíduos heterozigotos para os dois pares de genes ob-
tém-se:
a) 3 brancos: 1 preto
b) brancos: 3 pretos: 3 mesclados de marrom e preto: 1 branco
c) 1 preto: 2 brancos: 1 marrom
d) 12 brancos: 3 pretos: 1 marrom
e) 3 pretos: 1 branco
18 – Sobre os tipos sanguíneos do sistema ABO, assinale o que for correto.
a) Os indivíduos do tipo sanguíneo AB possuem aglutinina anti-A e anti-B
no plasma sanguíneo.
b) A herança genética do sistema ABO se processa por meio de quatro
pares de alelos, constituindo, portanto, um caso de polialelia.
c) O tipo sanguíneo O é muito frequente e, por este motivo, o alelo
responsável por sua expressão é dominante sobre os demais.
d) Os indivíduos do tipo sanguíneo O possuem aglutinogênios em suas
hemácias, porém não possuem aglutininas no plasma.
e) A transfusão de sangue de um doador do tipo A para um receptor do
tipo B provoca aglutinação, pois o receptor possui aglutinina anti-A
no plasma.
75.
74
19 – (UPE)Na síndrome de Waardenburg, os afetados apresentam deficiência
auditiva e discretas anomalias da face, além de modificação do pigmento
(pele, cabelo, olho). Diferentes membros de uma mesma família podem
exibir aspectos distintos da síndrome, podendo oscilar desde a perda
moderada de audição e mecha branca no cabelo até a surdez profunda,
acompanhada da heterocromia da íris (olho direito e esquerdo com
cores diferentes) e grisalhamento precoce do cabelo. Essa variabilidade,
manifestada desde o fenótipo mais leve ao mais grave, em diferentes
indivíduos, é denominada:
A) Dominância.
B) Epistasia.
C) Expressividade.
D) Penetrância.
E) Pleiotropia.
20 – Na planta maravilha não há dominância entre os alelos CB, que
condiciona a cor branca, e CV, que determina cor vermelha. A planta
heterozigota é rósea. Realizou-se um cruzamento de planta de flor
rósea com planta também de flor rósea e obtiveram-se 32 descenden-
tes. Qual o número esperado de descendentes brancas, vermelhas e
róseas, na progênie?
a) 8 brancas, 8 vermelhas e 16 róseas.
b) 24 vermelhas e 8 brancas.
c) 24 róseas e 8 vermelhas.
d) 16 róseas e 8 vermelhas.
e) 24 róseas, 4 vermelhas e 4 brancas.
76.
75
CAPÍTULO IV
MAPEAMENTO CROMOSSÔMICO
Mariade Mascena Diniz Maia e Hildson Dornelas Angelo da Silva
Introdução
Para melhor compreensão do tema ligação gênica, iniciaremos com
uma revisão sobre a base cromossômica dos Princípios Mendelianos
da Segregação e da Distribuição Independente.
Todos os organismos vivos possuem seqüências polinucleotídicas
que permitem a sua sobrevivência, capacidade de regeneração, du-
plicação e reprodução. O desenvolvimento destes mecanismos tem
sua base nestas seqüências, chamadas genes. Nos organismos cujo
cromossomo é único, como no caso das bactérias e alguns vírus, todos
os genes estão ligados, enquanto que nos indivíduos que possuem mais
de um cromossomo, os genes poderão estar ligados ou não.
Nos indivíduos diplóides, como milho, soja, feijão, arroz, Drosophi-
la que possuem, respectivamente, 20, 40, 24, 22 e 4 cromossomos,
muitos de seus genes estão no mesmo cromossomo, como muitos
estão em cromossomos separados.
O estudo dos genes independentes iniciou com os trabalhos de
Mendel enquanto que a ligação foi descoberta por volta de 1903 por
Sutton e mais tarde por Bateson e Punnet (1905). Mas foi somente
em 1910 que T. H. Morgan evidenciou o processo de ligação em cro-
mossomos de Drosophila.
Gregor Mendel (1822-1884) determinou dois princípios da trans-
missão genética:
1.Princípio da Segregação ( 1ª Lei de Mendel) diz que os organismos
diploides possuem dois alelos para qualquer característica e que
estes dois alelos se segregam (separam) quando os gametas são
formados na meiose, indo cada alelo para cada gameta.
2. Princípio da distribuição independente (2ª Lei de Mendel) diz que
os alelos de genes diferentes (loci diferentes) se distribuem inde-
pendentemente uns dos outros.
77.
76
É importante entenderque o princípio da segregação e o prin-
cípio da distribuição independente não se referem a dois processos
diferentes, ou seja, o princípio da distribuição independente é uma
extensão do princípio da segregação. A Teoria Cromossômica da
Hereditariedade (diz que os genes estão situados em cromossomos)
desenvolvida por Walter Sutton em 1900 possibilitou explicar estes
princípios (bem como exceções a eles) em termos do comportamento
meiótico dos cromossomos.
Segregação – Na primeira meiose os cromossomos homólogos se
pareiam (um homólogo vem da mãe e outro do pai). Na anáfase da
primeira meiose os cromossomos pareados se separam e se deslocam
para pólos opostos da célula. Um leva o alelo A e o outro o alelo a. A
separação física dos dois cromossomos segrega os alelos um do ou-
tro que irão residir em células-filhas diferentes. Este é o princípio da
segregação de Mendel que se baseia na separação dos cromossomos
homólogos durante a anáfase da primeira meiose (Figura 1).
Fonte:Adaptado de
Figura 1 – 1ª Lei de Mendel – Princípio da Segregação
Fonte:Adaptado de
78.
77
Princípio da distribuiçãoindependente
Este princípio da distribuição independente se baseia também na
separaçãoanafásica. Para compreendermelhor acorrelação, considere-
mos os genes em dois pares diferentes de cromossomos. Suponhamos
que o heterozigoto Aa Bb foi produzido no cruzamento de uma fêmea
AA BB com um macho aa bb. Considere que os dois genes estão em
cromossomos diferentes. Durante a prófase da meiose I, os cromos-
somos com os alelos A e a se pareiam, como o fazem os cromossomos
com os alelos B e b. Na metáfase, os dois pares tomam posições no
fuso meiótico para a separação anafásica seguinte. Como existem
dois pares de cromossomos, existem dois alinhamentos metafásicos:
A B
ou
A b
a b a B
Qualquer um destes alinhamentos pode ocorrer. Nesta demons-
tração o espaço separa pares diferentes de cromossomos e as barras
separam os membros homólogos de cada par. Durante a anáfase, os
alelos acima das barras irão se mover para um pólo, e os alelos abaixo
delas irão se mover para o outro. Quando ocorre a disjunção, há 50%
de chance de que os alelos A e B se movam juntos para o mesmo pólo
e uma chance de 50% de que se movam para pólos opostos. Analoga-
mente, há uma chance de 50% de que os alelos a e b se dirijam para o
mesmo pólo e de 50% de que se dirijam para pólos opostos. Ao final
da meiose, quando o número de cromossomos é finalmente reduzido,
metade dos gametas deverá conter uma combinação parental de alelos
(A B ou a b) e metade deverá conter uma nova combinação (A b ou
a B). No final, existirão quatro tipos de gametas, cada um sendo um
quarto do total (Figura 2). Esta igualdade de frequências gaméticas
é um resultado do comportamento independente dos dois pares
de cromossomos durante a meiose I. Podemos, portanto concluir,
que o princípio da distribuição independente de Mendel, refere-se à
79.
78
separação anafásica degenes em pares diferentes de cromossomos.
Ainda neste capítulo veremos que os genes situados no mesmo par de
cromossomos não se distribuem independentemente. Em vez disso,
como eles estão fisicamente ligados uns aos outros, tendem a ir juntos
através da meiose, violando o princípio da distribuição independente.
Conceitos Importantes: Os genes estão situados em cromossomos.
A disjunção dos cromossomos durante a meiose é responsável pela
segregação e distribuição independente dos genes.
Teoria Cromossômica da Hereditariedade
As primeiras evidências para esta teoria foram descobertas por
Thomas Hunt Morgan em 1910 a partir de estudos com mosca de
fruta, a Drosophila melanogaster. Mosca de frutas era adequada para
pesquisas genéticas devido ao seu pequeno tamanho, baixo número
de cromossomos, fácil criação em pequenos espaços de laboratório,
baixocustoparamanutençãodasculturas,geraçãocomgrandenúmero
74
distribuição independente.
Conceitos Importantes: Os genes estão situados em cromossomos. A
disjunção dos cromossomos durante a meiose é responsável pela segregação
e distribuição independente dos genes.
Teoria Cromossômica da Hereditariedade
As primeiras evidências para esta teoria foram descobertas por Thomas Hunt
Morgan em 1910 a partir de estudos com mosca de fruta, a Drosophila
melanogaster. Mosca de frutas era adequada para pesquisas genéticas devido
ao seu pequeno tamanho, baixo número de cromossomos,fácil criação em
pequenos espaços de laboratório, baixo custo para manutenção das culturas,
geração com grande número de descendentes em pouco espaço de tempo e
Figura 2 – 2ª Lei de Mendel – Princípio da Distribuíção Independente
80.
79
de descendentes empouco espaço de tempo e possibilidade de
obtenção de fêmeas virgens logo após a eclosão para efetuar cru-
zamentos recíprocos. Além disso, este inseto tem apenas quatro
pares de cromossomos, sendo um par de cromossomos sexuais,
XX nas fêmeas e XY nos machos. Morgan mostrou por meio de
experimentos, que a mutação de cor de olho dessas moscas era
herdada junto com o cromossomo X sugerindo que o gene para
essa característica estava fisicamente situado nesse cromossomo.
Entretanto, mais tarde, Calvin B. Bridges, um dos estudantes de
Morgan, obteve a prova definitiva da Teoria Cromossômica da
Hereditariedade.
Experimento de Bridges para comprovar que o gene para
olhos brancos da Drosophila está situado no cromossomo X
(Teoria Cromossômica da Herança).
Bridges cruzou uma fêmea de olhos brancos (Xw
Xw
) com um
macho de olhos vermelhos (X+
Y) e observou que cerca de 2,5% da
prole masculina tinham olhos vermelhos e 2,5% da prole feminina
tinham olhos brancos. No cruzamento está claro que cada mosca
masculina deveria herdar o cromossomo X da mãe ficando com o
genótipo Xw
Y ou seja macho de olhos brancos. As moscas fêmeas
do cruzamento deveriam herdar um alelo dominante para olhos
vermelhos no cromossomo X do pai juntamente com um alelo
para os olhos brancos no cromossomo X da mãe. A prole feminina
resultante, portanto deveria ser X+
Xw
, todas com olhos vermelhos.
A cor vermelha é dominante sobre a cor branca. Entretanto, o sur-
gimento de machos de olhos vermelhos e fêmeas de olhos brancos
nesse cruzamento foi inesperado. Bridges explicou esse resultado
formulando a hipótese de que, ocasionalmente, os dois cromossomos
X nas fêmeas poderiam não se separar durante a anáfase I da meio-
se. Essa falha na separação dos cromossomos Bridges chamou de
não-disjunção. Assim, alguns ovócitos poderão receber duas cópias
de X e outros não receberão nenhum X. Na fertilização poderão
81.
80
surgir os seguinteszigotos após a fertilização dos ovócitos com
os espermatozoides de machos com olhos vermelhos: ovócitos
XX (não-disjunção) fertilizado com espermatozoide portando Y
resulta em zigoto Xw
Xw
Y. Como o sexo da Drosophila é determi-
nado pela proporção X:A que é igual a 1,0 , logo o zigoto Xw
Xw
Y
é fêmea de olhos brancos. O ovócito de com dois cromossomo X
fertilizado com um espermatozoide portador de X produz o zigoto
Xw
Xw
X+
que morre. Um ovócito sem cromossomo X fertilizado
por um espermatozoide portador de X produz X+
O se desenvol-
ve em macho de olhos vermelhos estéreis e se um ovócito sem
cromossomo X é fertilizado por um espermatozoide portador de
Y resulta em zigoto YO sem cromossomo X e morre (Figura 3).
Bridges encontrou em seu experimento que a rara não disjunção
de cromossomos X entre as fêmeas de olhos brancos produz
alguns machos de olhos vermelhos e fêmeas de olhos brancos.
O significado do estudo de Bridges não foi ter explicado o surgi-
mento de uma mosca ocasional diferente em sua cultura, mas ser
capaz de prever a constituição cromossômica da Drosophila com
base em seu genótipo para cor de olho. Essa evidente associa-
ção entre genótipo e os cromossomos foi decisivo para mostrar
que os genes ligados ao sexo estão situados no cromossomo X,
confirmando a Teoria Cromossômica da Herança.
Ligação, Recombinação e Crossing over
Alfred Henry Sturtevant (1891 - 1970), geneticista americano
e colaborador de Thomas Morgan (1866-1945), desenvolveu o
primeiro mapa genético trabalhando com moscas e se baseando
no princípio de que os genes no mesmo cromossomo devem ser
herdados juntos porque estão fisicamente ligados na mesma estru-
tura e podem “viajar” como uma unidade durante a meiose. Este
fenômeno foi chamado de Ligaçâo. Ele reconheceu que poderia
usar a frequência de recombinação (FR) como uma medida da
distância linear entre dois genes. Os primeiros geneticistas sabiam
82.
81
que o processode ligação não era absoluto, de forma que, os genes
situados no mesmo cromossomo podiam ser separados quando
passassem pela meiose formando novas combinações de genes.
Fenômeno este, que foi chamado de Recombinação. O crossing over
que ocorre na prófase I da meiose é a troca de material genético
entre cromátides não irmãs de um par de cromossomo homólogo e
representa a base física da recombinação. As duas cromátides que
não participam do crossing over são chamadas parentais e formam
gametas não recombinantes. Enquanto que, as duas cromátides
que participam do crossing contém novas combinações de alelos
e formam gametas recombinantes (Figura 4).
Figura 3 – A Não-disjunção do cromossomo X é responsável pela prole excepcional
surgida no experimento de Bridges.
Figura 3. A Não-disjunção do cromossomo X é responsável pela
excepcional surgida no experimento de Bridges.
Ligação, Recombinação e Crossing over
Alfred Henry Sturtevant (1891 - 1970), geneticista america
colaborador de Thomas Morgan (1866-1945), desenvolveu o primeiro
genético trabalhando com moscas e se baseando no princípio de que os
no mesmo cromossomo devem ser herdados juntos porque estão fisica
83.
82
Durante a meiose,quando os cromossomos homólogos se
pareiam, uma troca física de material separa e recombina genes.
O fato é que os cromossomos nesse processo trocam pedaços
entre si. No ponto de troca os dois homólogos fazem um crossing
como se tivesse quebrado e se reunem com o seu parceiro. Um
ponto do crossing foi denominado quiasma que do grego significa
“cruzamento”. Os geneticistas, então, passaram a usar o termo
Crossing Over, para descrever o processo que criava os quiasmas,
ou seja, troca entre os cromossomos pareados. Assim, os gene-
ticistas, consideraram a recombinação (a separação de genes
ligados e a formação de novas combinações gênicas) como sendo
resultado do processo crossing over. A importância do crossing
over, portanto, é o aumento da variabilidade genética.
Figura 4 – Um único crossing over produz metade de gametas não recombinantes e
metade recombinante.
78
participam do crossing contém novas combinações de alelos e formam
gametas recombinantes (Figura 4).
.
Figura 4. Um único crossing over produz metade de gametas não
recombinantes e metade recombinante.
Durante a meiose, quando os cromossomos homólogos se pareiam,
uma troca física de material separa e recombina genes. O fato é que os
cromossomos nesse processo trocam pedaços entre si. No ponto de troca os
dois homólogos fazem um crossing como se tivesse quebrado e se reunem
com o seu parceiro. Um ponto do crossing foi denominado quiasma que do
grego significa “cruzamento”. Os geneticistas, então, passaram a usar o termo
CROSSING OVER, para descrever o processo que criava os quiasmas, ou
a, troca entre os cromossomos pareados. Assim, os geneticistas,
sideraram a recombinação (a separação de genes ligados e a formação de
as combinações gênicas) como sendo resultado do processo crossing over.
mportância do crossing over, portanto, é o aumento da variabilidade
ética.
Figura 5 Formação de quiasma entre cromátides não irmãs de um par de
cromossomos homólogos.
Figura 5 – Formação de quiasma entre cromátides não irmãs
de um par de cromossomos homólogos.
84.
83
Conceitos Importantes: Ataxa de crossing entre dois genes ligados
indica a distância entre eles: alta taxa significa genes muito afastados,
baixas taxas significam genes muito próximos.
Ligação e Recombinação entre dois genes
Agora que temos a noção clara dos conceitos de ligação, recom-
binação e crossing over, passemos a detalhar melhor aplicando o que
aprendemos.
Iniciaremos estudando características genéticas cujos genes não
segregam independentemente. Como sabemos, este fenômeno se
refere à ligação gênica ou linkage, e foi descoberto em 1905 pelos
pesquisadores: William Bateson, Edith Rebecca Saunders e Reginald
C. Punnett que verificaram a falta de independência de dois genes de
ervilha. Estes genes não seguiam a Lei da Segregação Independente
(2ª Lei de Mendel). Assim, quando esses pesquisadores cruzaram uma
linhagem homozigota de ervilha com flores vermelhas (R) e grãos de
pólen longos (L) com uma linhagem homozigota com flores brancas
(r) e grãos de pólen pequeno (l) obtiveram a F1 com flores vermelhas
e grãos de pólen longos indicando que o vermelho era dominante em
relação ao branco e longo era dominante em relação a pequeno.
Entretanto, quando entrecruzaram a F1, a F2 resultante não apa-
recia na proporção 9:3:3:1 esperada na distribuição independente. Em
vez disso, aparecia um excesso de plantas F2 com flores vermelhas e
pólen longo ou flores brancas e pólen pequeno (fenótipos parentais
não recombinantes). O que estaria ocorrendo? Note que as duas classes
fenotípicas, vermelho longo e branco pequeno, são maiores do que o
esperado. Os pesquisadores observaram que a F1 tinha produzido mais
gametas parentais R.L e r.l do que seria produzido pela segregação
mendeliana independente. Haveria um acoplamento físico (proximi-
dade física) entre os alelos dominantes R e L e entre os recessivos r
e l impedindo sua distribuição independente na F1? Esta hipótese de
Bateson e Punnett sobre o acoplamento físico só pode ser desvendada
após os experimentos de Thomas Hunt Morgan que comprovaram o
85.
84
acoplamento de doisgenes autossômicos de Drosophila melanogaster,
um para cor de olho e outro para tamanho da asa. .
A pergunta é: porque usar genes “ligados” em vez de ”acoplados”?
A resposta é que as palavras acoplamento e repulsão são usadas para
identificar dois tipos diferentes de conformação de ligação em um
heterozigoto duplo:
Conformação acoplada ou CIS
Conformação em repulsão ou TRANS
O acoplamento refere-se à ligação de dois alelos dominantes ou
dois recessivos e a repulsão indica que os alelos dominantes estão
ligados a alelos recessivos. Para avaliar se um duplo heterozigoto está
em acoplamento ou repulsão deve-se fazer um cruzamento teste com
o duplo heterozigoto, ou considerar os genótipos de seus genitores.
Conceitos Importantes: A recombinação genética é uma fonte de
variabilidade genética (evolução), ocorre em espécies que se repro-
duzem sexualmente e pode viabilizar a junção de alelos favoráveis de
genes diferentes no mesmo indivíduo.
Cálculo da Frequência de Recombinação
Na herança de genes ligados a frequência dos gametas de um
heterozigoto depende da taxa de crossing over (ou taxa de recom-
binação) que ocorre entre os cromossomos homólogos. Os gametas
parentais são formados mesmo que não haja recombinação e apare-
ce em maior quantidade. Enquanto os gametas recombinantes são
formados apenas quando houver permutas e aparecem em menor
quantidade. A taxa ou frequência de crossing over entre dois genes
que as palavras acoplamento e repulsão são usadas para identificar dois
os diferentes de conformação de ligação em um heterozigoto duplo:
onformação acoplada ou CIS
onformação em repulsão ou TRANS
O acoplamento refere-se à ligação de dois alelos dominantes ou dois
cessivos e a repulsão indica que os alelos dominantes estão ligados a alelos
cessivos. Para avaliar se um duplo heterozigoto está em acoplamento ou
pulsão deve-se fazer um cruzamento teste com o duplo heterozigoto, ou
nsiderar os genótipos de seus genitores.
onceitos Importantes
A recombinação genética é uma fonte de variabilidade genética
volução), ocorre em espécies que se reproduzem sexualmente e pode
abilizar a junção de alelos favoráveis de genes diferentes no mesmo
divíduo.
que as palavras acoplamento e repulsão são usadas para identificar dois
os diferentes de conformação de ligação em um heterozigoto duplo:
nformação acoplada ou CIS
nformação em repulsão ou TRANS
O acoplamento refere-se à ligação de dois alelos dominantes ou dois
cessivos e a repulsão indica que os alelos dominantes estão ligados a alelos
cessivos. Para avaliar se um duplo heterozigoto está em acoplamento ou
pulsão deve-se fazer um cruzamento teste com o duplo heterozigoto, ou
nsiderar os genótipos de seus genitores.
onceitos Importantes
A recombinação genética é uma fonte de variabilidade genética
volução), ocorre em espécies que se reproduzem sexualmente e pode
bilizar a junção de alelos favoráveis de genes diferentes no mesmo
divíduo.
86.
85
é definida comoa soma das frequências em que ocorrem os gametas
recombinantes. A frequência da prole recombinante (aquela com
fenótipo diferente de ambos os pais) pode ser calcula. Podemos usar
a frequência de recombinação para medir a intensidade de ligação. O
modo mais direto de estimar a frequência de recombinação (FR) é usar
umcruzamento-testeonde umindivíduoaseranalisadogeneticamente
é cruzado com um homozigoto que possui alelos recessivos. A Figura 6
abaixo mostra um cruzamento-teste da F1
resultante do cruzamento
entre ervilhas visto na seção anterior. A prole deste cruzamento-teste
revela os tipos de gametas e as proporções produzidas nas plantas
heterozigotas da F1
.
83
Para quaisquer dois genes ligados, a frequência de gametas recombinantes
nunca excede a 50%. Podendo atingir esse valor se os genes situados nos
mesmos cromossomos estiverem muito distantes (nas pontas opostas de um
cromossomo). Este valor é também atingido quando os genes estão em
cromossomos diferentes (diibridismo, 2ª Lei de Mendel): 50% de recom
binantes significa que os genes se distribuem
independentemente.
Figura 6 – Cruzamento-teste para ligação entre genes de ervilha. como a prole
recombinante na F é igual a 8% do total os genes para cor de flores e tamanho do
pólen estão proximamente ligados
87.
86
Analisando os dadosdo cruzamento-teste podemos observar que
foram produzidas 1000 indivíduos sendo, 920 de uma ou outra linha-
gem parental e 80 indivíduos recombinantes. O cálculo da Frequência
de Recombinação (FR) pode ser feito usando a seguinte fórmula:
FR=
∑ recombinante
x 100%
Número total da prole
Com o uso desta fórmula acha-se a distância entre os genes em
estudo. Os resultados indicam que a “frequência de recombinação
é diretamente proporcional à distância entre os genes”.
Para quaisquer dois genes ligados, a frequência de gametas
recombinantes nunca excede a 50%. Podendo atingir esse valor
se os genes situados nos mesmos cromossomos estiverem muito
distantes (nas pontas opostas de um cromossomo). Este valor é
também atingido quando os genes estão em cromossomos diferen-
tes (diibridismo, 2ª Lei de Mendel): 50% de recombinantes significa
que os genes se distribuem independentemente.
Portanto, o cálculo acima resultou em 8% de recombinação isso
significa que o gene “l” dista de “r” em 8 unidades de mapa ou 8 cM
(centi Morgan). A frequência baixa de recombinação indica que os
genes estão bem ligados.
Mapeamento gênico em eucarioto
Mapeamento genético ou cromossômico é a representação da
posição dos genes no cromossomo e tem como objetivo analisar a
frequência de recombinação alélica em cruzamentos.
Novas combinações de alelos parentais são produzidas por
recombinação (“crossing over” durante a meiose I) e cruzamentos-
teste são usados para determinar quais genes estão ligados e criar
um mapa de ligação (mapa genético de cada cromossomo). Morgan e
seus colaboradores haviam determinado as frequências de recombi-
88.
87
nação para muitospares de genes ligados de Drosophilas. Sturtevant
utilizou estes dados para criar mapas genéticos que mostravam o
arranjo de genes ao longo do cromossomo. A partir do método de
Sturtevant, os geneticistas têm mapeado os cromossomos de vários
organismos eucariotos, procariotos e vírus indicando as distâncias
entre genes em unidade de mapa (u.m.). Por definição, uma unidade
de mapa corresponde a uma frequência de recombinação de 0,01
também referida como um centiMorgan (cM) em homenagem a
Morgan. 100 centiMorgan são iguais a um Morgan.
Recombinação com cruzamento-teste de dois pontos
A Figura 7 ilustra um cruzamento teste de dois pontos onde fê-
meas do tipo selvagem de Drosophila (vg+ e b+ asas longas e corpo
cinza respectivamente) foram cruzadas com machos homozigotos
para duas mutações autossômicas, asas vestigiais (vg) e black (b)
asas pequenas e corpo preto. Na F1
, foi observado que todas as
moscas tinham asas longas e corpo cinza, logo se deduziu que os
alelos tipos selvagem vg+ e b+ são dominantes.
As fêmeas F1
(vgvg+ bb+) foram submetidas ao cruzamento-
teste com machos vestigiais pretos(vgvg bb) e a prole F2
resultante,
foi distribuída e contada. Os dados mostram quatro classes feno-
típicas: as parentais, mais abundantes, com o mesmo fenótipo dos
genitores originais e as classes raras com fenótipos recombinantes.
Evidentemente, os genes para os fenótipos vestigial e black estão
ligados porque a classe de recombinantes aparece bem menos que
50% do total da prole indicando que estes genes estão no mesmo
cromossomo.
Para determinar a distância entre os genes vg e b calcula-se o
número médio de crossings nos gametas das fêmeas F1
duplo-he-
terozigotas. Para isto, calculamos a frequência de moscas recom-
binantes F2
observando que cada mosca herdou um cromossomo
que tinha um crossing entre vg e b
89.
88
Número médio= nãorecombinante (0) x (0,82) + recombinantes (1)
x0,18 = 0,18.
Observe que 0,82 é resultado de 100- 0,18 (frequência de recombi-
nação).
Figura 7 – Experimento envolvendo dois genes ligados,
vg (asas vestigiais) e b (corpo black ) em Drosophila.
90.
89
A prole nãorecombinante não soma nenhum cromossomo com
crossing aos dados, mas deve ser incluída no cálculo para enfatizar que
devemos calcular o número médio de crossing usando todos os dados,
e não apenas os dos recombinantes. A frequência de recombinação de
0,18 indica que em média, 18 dos 100 cromossomos recuperados da
meiose tinham crossings entre vg e b. Concluímos então, que vg e b
estão separados por 18 unidades de mapa (u.m) ou 18 cM (ou 0,18 M).
Construção do mapa gênico:
Parentais 0820
Recombinantes 0180
Total 1000
FR = 180÷1000 = 0,18×100 = 18 u.m.
Mapa gênico:
18 u.m
vg b
Mapeamento de recombinação
em cruzamento-teste de três pontos
Para fazer esse mapeamento consideremos a figura 8 que ilustra
um cruzamento de três genes ligados realizado por Bridges e Olbrycht.
Eles cruzaram machos tipo selvagem de Drosophila com fêmeas homo-
zigotas para três mutações recessivas ligadas ao X: cerdas scute (sc),
olhos echinus (ec) e asas crossveinless(cv). Entrecruzaram a F1
e pro-
duziram moscas F2
, classificaram e contaram. As moscas fêmeas F1
neste cruzamento levaram as três mutações recessivas em um dos
seus cromossomos X e os alelos tipo selvagem destas mutações em
outro cromossomo X e os machos de F1
levaram as três mutações
recessivas em seu único cromossomo X. Deste entrecruzamento
91.
90
resultou uma proleF2
com oito classes fenotípicas distintas, duas
delas parentais e seis recombinantes. Sendo que as classes parentais
são mais numerosas. As classes recombinantes, menos numerosas,
representam cada tipo diferente de cromossomo com crossing. Assim,
para entender quais crossings estão envolvidos na produção de cada
tipo de recombinante, devemos primeiro determinar como os genes
estão ordenados no cromossomo.
representam cada tipo diferente de cromossomo com crossing.
entender quais crossings estão envolvidos na produção de
recombinante, devemos primeiro determinar como os genes estã
no cromossomo.
Figura 8. Cruzamento de três pontos de Bridges e Olbrycht com g
a X, sc (cerdas scute) e ec (olhos echinus) e cv (sas cross
Figura 8 – Cruzamento de três pontos de Bridges e Olbrycht com genes ligados
a X, sc (cerdas scute) e ec (olhos echinus) e cv (sas crossveinless) em Drosophila.
92.
91
Determinação da ordemdos genes no cromossomo
Imagine que pode haver, neste exemplo, três possíveis ordens de
genes:
1. sc – ec – cv
2. ec – sc – cv
3. ec – cv – sv
Qual das ordens é a correta?
1. sc- ec- cv
2. ec- sc- cv
3. ec- cv- sv
Qual das ordens é a correta?
Figura 9. Formação de recombinantes e cálculos da distância de mapa
genético dos dados de Bridges e Olbrycht.
Observe na Figura 9 que temos 8 classes assim distribuídas:
Parentais:
Figura 9 – Formação de recombinantes e cálculos da distância de mapa
genético dos dados de Bridges e Olbrycht.
Observe na Figura 9 que temos 8 classes assim distribuídas:
Parentais:
1. sc ec cv
2. sc+ ec+ cv+
93.
92
Recombinantes simples (envolvendo1 crossing) entre sc e ec:
3. sc ec+ cv+
4. sc+ ec cv
Recombinantes simples (envolvendo 1 crossing) entre ec e cv:
5. sc ec cv+
6. sc+ ec+ cv
Recombinantes duplos (envolvendo 2 crossings) 1 entre sc e ec e
outro entre ec e cv:
7. sc ec+ cv
8. sc+ ec cv+
Para determinar a ordem dos genes no cromossomo compare as
classes parentais com as classes duplos recombinantes. Nos duplos
recombinantes quem troca é o gene do meio. Assim, comparando
as classes parentais (1). sc ec cv e (2). sc+ ec+ cv+ com os duplos
recombinantes (7). sc ec+ cv e (8). sc+ ec cv+ vemos que o alelo echi-
nos (ec) foi substituído com relação a scute (sc) e crossveinless (cv).
Assim, podemos concluir que a ordem dos genes no cromossomo é:
1. sc – ec – cv.
Cálculo das distâncias entre os genes
Uma vez, estabelecido a ordem dos genes no cromossomo, pode-
mos agora determinar a distância entre os genes adjacentes.
Para obtenção do tamanho da região entre sc e ec, identificamos
as classes recombinantes que envolvem um crossing nesta região
incluindo as classes duplo recombinantes. Assim temos:
163+130+1+1 = 295÷3.248 = 0,091×100=9,1
isto significa que em cada 100 cromossomos vindos da meiose nas
fêmeas de F1
,9,1 tinha um crossing entre sc e ec. A Distância entre estes
94.
93
genes é 9,1u.m. ou 9,1 Centi Morgan. Da mesma maneira obtemos a
distância entre ec e cv. Temos:
192+148+1+1 = 342÷ 3.248 = 0,105×100 =
10,5 u.m. ou 10,5 CentiMorgan.
A distância entre sc e cv é a soma dos dois intervalos de mapa:
9,1cM +10,5 cM= 19,6cM.
Mapa Gênico:
sc ec cv
9,1 10,5
95.
94
CAPÍTULO V
DETERMINAÇÃO DOSEXO
Martín Alejandro Montes e Ana Cristina Lauer Garcia
Uma das características mais importantes dos organismos vivos é
a sua capacidade de se transmitir as informações genéticas através das
gerações. Existem dois tipos de reprodução, a assexuada e a sexuada
(Figura 1). Na reprodução assexuada, um indivíduo pode se dividir em
dois, deixando descendentes sem a necessidade de um parceiro, é o
caso dos procariotos ou bactérias e de alguns eucariotos. Os indiví-
duos que surgem por esta forma de reprodução são geneticamente
idênticos entre si, formando o que se chama de clone.
Figura 1 – As leveduras (A) são exemplos de organismos que se reproduzem
de forma assexuada, ou seja, sem a formação de gametas. Nas moscas (B) a
reprodução é sexuada, na qual ocorre a fusão de gametas masculinos e femininos,
formando novos indivíduos. A reprodução sexuada contribui para a variabilidade
genética das espécies.
A grande maioria dos organismos se reproduz de forma sexuada,
um processo que envolve a formação de gametas e a fusão destas
células, originando um novo indivíduo. O sexo permite a geração de
filhos geneticamente diferentes de seus progenitores e isso pode
representar uma vantagem adaptativa para as populações e espécies.
92
Martín Alejandro Montes e Ana Cristina Lauer Garcia
Uma das características mais importantes dos organismos vivos é a sua
capacidade de se transmitir as informações genéticas através das gerações.
Existem dois tipos de reprodução, a assexuada e a sexuada (Figura 1). Na
reprodução assexuada, um indivíduo pode se dividir em dois, deixando
descendentes sem a necessidade de um parceiro, é o caso dos procariotos ou
bactérias e de alguns eucariotos. Os indivíduos que surgem por esta forma de
reprodução são geneticamente idênticos entre si, formando o que se chama de
clone.
A grande maioria dos organismos se reproduz de forma sexuada, um
processo que envolve a formação de gametas e a fusão destas células,
originando um novo indivíduo. O sexo permite a geração de filhos
Figura 1. As leveduras (A) são exemplos de organismos que se
reproduzem de forma assexuada, ou seja, sem a formação de
gametas. Nas moscas (B) a reprodução é sexuada, na qual ocorre a
fusão de gametas masculinos e femininos, formando novos
indivíduos. A reprodução sexuada contribui para a variabilidade
genética das espécies.
96.
95
Os organismos dereprodução sexuada podem ser monoicos (do
grego mono=um e oikos=casa) quando um único indivíduo apresen-
ta dois sexos, ou dioicos (do grego di=duas e oikos=casa), quando o
organismo apresenta apenas um sexo, masculino ou feminino.
No caso dos organismos monoicos, também chamados de herma-
froditas, há exemplos de seres vivos, tal como a ervilha, em que o óvulo
e o espermatozoide utilizado para formar o novo ser são produzidos
pelo mesmo indivíduo, ocorrendo o que chamamos de autofecunda-
ção. Por outro lado, às vezes um único organismo apresenta os dois
sexos, mas não realiza autofecundação, como é o caso das minhocas
(Figura 2). Ainda entre os hermafroditas, há casos em que um indiví-
duo pode apresentar os dois sexos funcionais no mesmo momento
de sua vida ou não. Nesta situação estamos diante de um caso de
hermafroditismo sequencial. Exemplos de hermafroditas sequenciais
são representados pelo peixe-palhaço Amphiprion percula, no qual
o primeiro sexo a aparecer é o macho (um hermafrodita sequencial
protândrico) e pelo peixe Thalassoma lunare, em que a fêmea é o
sexo inicial (um hermafrodita sequencial protogínico).
Neste capítulo falaremos sobre como é determinado o sexo nos
organismos com reprodução sexuada.
ferentes de seus
isso pode
ma vantagem
as populações e
ganismos de
ada podem ser
ego mono=um e
ando um único
nta dois sexos,
grego di=duas e
ndo o organismo
nas um sexo,
minino.
dos organismos monoicos, também chamados de
á exemplos de seres vivos, tal como a ervilha, em que o óvulo
oide utilizado para formar o novo ser são produzidos pelo
, ocorrendo o que chamamos de autofecundação. Por outro
m único organismo apresenta os dois sexos, mas não realiza
como é o caso das minhocas (Figura 2). Ainda entre os
á casos em que um indivíduo pode apresentar os dois sexos
esmo momento de sua vida ou não. Nesta situação estamos
so de hermafroditismo sequencial. Exemplos de hermafroditas
representados pelo peixe-palhaço Amphiprion percula, no qual
aparecer é o macho (um hermafrodita sequencial protândrico)
halassoma lunare, em que a fêmea é o sexo inicial (um
uencial protogínico).
Figura 2. A minhoca é um exemplo de
organismo hermafrodita. Neste caso, um
único indivíduo apresenta os dois sexos,
masculino e feminino, mas a reprodução se
dá pela cópula, já que não ocorre
autofecundação.
Figura 2 – A minhoca é um exemplo de organismo hermafrodita. Neste caso, um
único indivíduo apresenta os dois sexos, masculino e feminino, mas a reprodução se
dá pela cópula, já que não ocorre autofecundação.
97.
96
Mecanismos de Determinaçãodo Sexo
A determinação do sexo sempre depende de uma base genética.
Em alguns organismos a regulação dos genes responsáveis pela defi-
nição sexual pode ser muito influenciada pelo ambiente. Em outros,
é a quantidade de cromossomos que apresenta um papel decisivo na
determinação do ser masculino ou feminino. Há casos também de
organismos que apresentam um conjunto limitado de cromossomos,
geralmente dois, responsáveis pela definição do sexo. Deste modo,
existem dois grandes sistemas de determinação do sexo: o ambiental
e o cromossômico.
Sistemas de Determinação Ambiental do Sexo
Existem vários exemplos de determinação ambiental do sexo. Neste
sistema, a concentração, ou o tipo de substâncias no ambiente, assim
como a temperatura são responsáveis por ativar ou desligar deter-
minados genes no organismo, o que garante o seu desenvolvimento
como um macho ou como uma fêmea.
Um exemplo de determinação sexual em que substâncias liberadas
pelo ambiente são decisivas para a formação do sexo masculino ou
feminino é ilustrada pelo molusco marinho Crepidula fornicata. Este
organismo tem uma fase juvenil larval e uma fase adulta séssil. Quando
uma larva encontra um local longe de outros moluscos de sua espécie
esta se fixa em um substrato e se desenvolve como uma fêmea. Essa
fêmea produz substâncias, responsáveis pela atração de outras larvas
de sua espécie, as quais se fixam perto dela e se desenvolvem como
machos. O primeiro indivíduo garante, desta forma, os parceiros para
sua reprodução. Após um tempo, os machos transformam-se em
fêmeas, liberando mais substâncias que atraem outras larvas que, ao
se fixarem sobre estas, viram machos. A transformação dos machos
em fêmeas é conhecida como hermafroditismo sequencial.
Outro exemplo de determinação ambiental do sexo é observado no
verme marinho Bonellia virides. Este se apresenta inicialmente como
uma larva indiferenciada sexualmente. Caso a larva não encontre
98.
97
Nos crocodilos eem algumas espécies de tartarugas e lagartos,
do filhote é determinado pela temperatura ambiental duran
desenvolvimento do embrião (Figura 3). Existem três situaçõe
determinação do sexo pela temperatura, há casos, como no lagarto t
(Sphenodon punctatus), em que temperaturas altas determina
desenvolvimento de machos. Em outros casos, temperaturas mais ele
levam a diferenciação de fêmeas, como exemplificado com as tartarug
espécie Chrysemys picta. Finalmente, temperaturas altas e baixas
produzir fêmeas e temperaturas intermediárias levar a formação de m
este é caso dos crocodilianos. O valor da temperatura que limita o sexo
variar de acordo com a espécie.
Figura 3. Em algumas espécies de
lagartos, tal como Sphenodon punctatus
(A), de tartarugas, tal como Chrysemys
picta (B) e em crocodilianos (C), a
determinação do sexo ocorre pela
ativação de genes diferenciais
dependendo da temperatura de
desenvolvimento dos embriões.
uma fêmea próxima no momento da diferenciação sexual esta se
diferenciará em fêmea. Porém se a larva encontrar uma fêmea por
perto se diferenciará em macho. Tal situação é determinada por
uma substância produzida pela fêmea adulta chamada de bonellin.
O macho vive dentro do corpo da fêmea, garantindo a produção
de esperma para a reprodução.
Nos crocodilos e em algumas espécies de tartarugas e lagar-
tos, o sexo do filhote é determinado pela temperatura ambiental
durante o desenvolvimento do embrião (Figura 3). Existem três
situações de determinação do sexo pela temperatura, há casos,
como no lagarto tuatara (Sphenodon punctatus), em que tempera-
turas altas determinam o desenvolvimento de machos. Em outros
casos, temperaturas mais elevadas levam a diferenciação de fêmeas,
como exemplificado com as tartarugas da espécie Chrysemys picta.
Finalmente, temperaturas altas e baixas podem produzir fêmeas e
temperaturas intermediárias levar a formação de machos, este é
caso dos crocodilianos. O valor da temperatura que limita o sexo
pode variar de acordo com a espécie.
Nos crocodilos e em algumas espécies de tartarugas e lagartos, o sexo
do filhote é determinado pela temperatura ambiental durante o
desenvolvimento do embrião (Figura 3). Existem três situações de
determinação do sexo pela temperatura, há casos, como no lagarto tuatara
(Sphenodon punctatus), em que temperaturas altas determinam o
desenvolvimento de machos. Em outros casos, temperaturas mais elevadas
levam a diferenciação de fêmeas, como exemplificado com as tartarugas da
espécie Chrysemys picta. Finalmente, temperaturas altas e baixas podem
produzir fêmeas e temperaturas intermediárias levar a formação de machos,
este é caso dos crocodilianos. O valor da temperatura que limita o sexo pode
variar de acordo com a espécie.
SISTEMA HAPLOIDE/DIPLOIDE DE DETERMINAÇÃO DO SEXO
Figura 3. Em algumas espécies de
lagartos, tal como Sphenodon punctatus
(A), de tartarugas, tal como Chrysemys
picta (B) e em crocodilianos (C), a
determinação do sexo ocorre pela
ativação de genes diferenciais
dependendo da temperatura de
desenvolvimento dos embriões.
Figura 3 – Em algumas espécies
de lagartos, tal como Sphenodon
punctatus (A), de tartarugas, tal como
Chrysemys picta (B) e em crocodilianos
(C), a determinação do sexo ocorre
pela ativação de genes diferenciais
dependendo da temperatura de
desenvolvimento dos embriões.
99.
98
Sistema Haploide/Diploide deDeterminação do Sexo
Em alguns insetos como abe-
lhas,vespaseformigasosistemade
determinação do sexo é um tanto
peculiar, já que envolve o conjunto
completo de cromossomos. Neste
caso as fêmeas apresentam dois
conjuntoscromossômicos(2n)eos
machos apenas um (n). Por meiose
as fêmeas produzem gametas com
um conjunto cromossômico (n). Os
machos também formam gametas
com um conjunto cromossômico,
porém isto ocorre por mitose. A fe-
cundação de um gameta feminino
(n) com um gameta masculino (n)
forma fêmeas diploides (2n).
A partir de ovos não-fecunda-
dos ocorre a formação dos machos
(no caso das abelhas, os zangões)
por um processo conhecido como
partenogênese. Os machos apre-
sentam um único conjunto cro-
mossômico totalmente herdado
da mãe e são, portanto, haploides
(n). A Figura 4 ilustra este sistema
de determinação do sexo.
Sistema de Determinação do Sexo pela Relação
entre Cromossomos Sexuais e Não Sexuais
No caso das moscas das frutas, as drosófilas, a determinação do
sexo depende da razão entre o número de cromossomos sexuais (X)
e o número de conjuntos haploides de autossomos (A), ou simples-
mitose. A
feminino (n
(n) forma fê
A pa
ocorre a for
das abelh
processo
partenogên
um único
totalmente
portanto, ha
este sistem
SISTEMA DE DETERMINAÇÃO DO SEXO
CROMOSSOMOS SEXUAIS E NÃO SEXUAIS
No caso das moscas das frutas, as drosófi
depende da razão entre o número de cromossomo
conjuntos haploides de autossomos (A), ou simpl
tipo de determinação de sexo é chamado de Siste
As fêmeas normais possuem dois cromo
haploides de autossomos, sendo a razão X/A ig
possuem apenas um cromossomo X para
autossômicos, numa razão de 0,5. Indivíduos que
que um são denominados de metafêmeas, enquan
Figura 4. N
determinação
dá pelo
cromossômic
cromossômic
fêmeas e
machos.
Figura 4 – No sistema haploide/
diploide de determinação do sexo a
definição sexual se dá pelo número
de conjuntos cromossômicos. Dois
conjuntos cromossômicos levam à
formação de fêmeas e um conjunto à
formação de machos.
100.
99
mente da razãoX/A. Esse tipo de determinação de sexo é chamado de
Sistema de balanço gênico.
As fêmeas normais possuem dois cromossomos X e dois conjuntos
haploides de autossomos, sendo a razão X/A igual a um. Machos nor-
mais possuem apenas um cromossomo X para dois conjuntos haploides
autossômicos, numa razão de 0,5. Indivíduos que possuem uma razão
maior que um são denominados de metafêmeas, enquanto indivíduos
com uma razão menor do que 0,5 são definidos como metamachos.
Já indivíduos com razão entre 0,5 e 1 apresentam um fenótipo inter-
mediário chamado de intersexo.
Sistemas de Determinação Cromossômica do Sexo
Em alguns organismos os genes principais para a determinação
do sexo se encontram em um grupo de cromossomos, geralmente
um par, chamados de cromossomos sexuais ou alossomos. Os demais
cromossomos são chamados de autossomos.
Em algumas espécies os cromossomos que formam o par sexual
podem ser morfologicamente iguais entre si em ambos os sexos, exem-
plos deste caso são ilustrados por algumas populações do anfíbio Rana
rugosa e do peixe Apareiodon piracicabae. Nestes casos, é muito difícil
de visualizar os cromossomos sexuais em preparações citológicas
convencionais.
Na grande maioria dos casos, no entanto, os cromossomos sexuais
diferem morfologicamente entre si. Em muitas espécies um dos sexos
apresentacromossomossexuaismorfologicamenteidênticosenooutro
sexo o par sexual é diferente entre si. Tal situação permite definir dois
grandes sistemas de determinação do sexo, aquele em que o macho
apresenta os cromossomos sexuais diferentes entre si (sistemas XX
-XY e XX-X0) e aquele em que é a fêmea que apresenta esta condição
(sistemas ZZ-ZW e ZZ-Z0).
Nos sistemas de determinação do sexo denominados de XX-XY e
XX-X0 o macho é responsável pela definição do sexo da progênie. No
primeiro caso, os machos (XY) produzem metade dos gametas com
101.
100
o cromossomo Xe a outra metade com o cromossomo Y. Já a
fêmea (XX) produz apenas gametas com o cromossomo X. Caso o
gameta feminino seja fecundado por um X do pai é formada uma
fêmea e, no caso da junção de um X materno com um Y paterno
há a formação de um macho. Como o macho pode formar dois
tipos de gametas este é considerado o sexo heterogamético. No
caso do sistema XX-XY, tanto a fêmea como o macho apresen-
tam o mesmo número cromossômico. O sistema XX-XY ocorre
na maioria dos vertebrados, em mamíferos, em alguns insetos,
diversos tipos de peixes e em algumas plantas.
No sistema XX-X0 as fêmeas são XX enquanto os machos são
X0. Nesse caso, o “zero” representa a falta de um cromossomo
sexual. Assim como no sistema XX-XY, os machos seguem sen-
do o sexo heterogamético, já que metade dos espermatozoides
apresenta um cromossomo X e a outra metade não carrega este
cromossomo. Novamente cabe aos machos a determinação do
sexo da prole. O sistema XX-X0 ocorre em algumas aranhas e
muitos insetos da ordem dos odonatas, ortópteros, hemípteros
e heterópteros.
Nos sistemas de determinação do sexo denominados de ZZ-
ZW e ZZ-Z0 a fêmea é o sexo heterogamético. Nestes sistemas de
determinação do sexo os cromossomos sexuais recebem nomes
diferentes (Z e W) para evitar confusão com o sistema XX/XY e
XX/XO descritos anteriormente.
O sistema ZZ-ZW ocorre em muitas espécies de aves, répteis
e lepidópteros (mariposas e borboletas), neste caso os machos
apresentam o par de cromossomos sexuais homólogos (ZZ),
enquanto as fêmeas possuem um par não-homólogo (ZW).
No sistema ZZ-ZO as fêmeas (Z0) possuem apenas um cromos-
somo sexual, sendo a ausência do cromossomo W indicada pelo
“zero”. Os machos apresentam o par de cromossomos sexuais ZZ.
Este sistema de determinação do sexo está presente em algumas
espécies de mariposas e alguns outros insetos.
102.
101
Determinação do Sexoem Humanos
Em humanos e nos mamíferos placentários que apresentam um sis-
tema de determinação do sexo do tipo XX-XY os cromossomos X e Y
possuemmorfologiaetamanhoscontrastantes,sendooYmenordoque
o X. Mesmo com essas diferenças, estes cromossomos pareiam durante
a meiose. O pareamento se dá pelo fato de apresentarem regiões com o
mesmoconteúdoinformativo,chamadasderegiõespseudoautossômicas,
as quais são observadas nos extremos dos cromossomos X e Y.
Neste grupo de organismos o sexo é determinado pelo cromossomo
Y. A evidência para este fato se dá pela observação de que indivíduos X0
são fêmeas, enquanto os XXY são machos.
A expressão de genes presentes no cromossomo Y induz a gônada
primordialasetransformaremtestículo.Oconjuntogênicoresponsável
pela determinação do sexo é grande e ainda tem muito a ser desco-
berto e compreendido. Existem mais de cinco genes relacionados ao
começo da determinação do sexo (WNT4, DAX1, SF1, WT1, SOX9 e
SRY), como exemplo descreveremos um deles, a fim de entender a
complexidade dos mecanismos.
NobraçocurtodocromossomoY,foradasregiõespseudoautossômi-
cas,éencontradoogeneSRY,umdosprincipaisresponsáveispeladetermi-
naçãodosexo.OgeneSRYcodificaparaofatordedeterminaçãotesticular,
sendosuaexpressãoresponsávelpeladefiniçãodosexomasculino.Ogene
SRYfoidescobertonoestudodehomensXXqueapresentavamumacópia
incomum deste gene em um dos cromossomos X e de mulheres XY, nas
quais o cromossomo Y apresentava uma deleção deste gene.
QuandoogeneSRYestápresenteefuncional,suaexpressãoestimula
ameduladagônadaembrionáriaasediferenciaremtestículo.Aprodução
detestosteronapelotestículoinduzoaparecimentodascaracterísticas
masculinas no indivíduo. A ausência deste gene, como ocorre normal-
mentenasmulheres,implicanafaltadofatordeterminantedetestículos,
provocando o desenvolvimento do córtex da gônada primitiva, o que
origina o ovário. Na ausência de grandes quantidades de testosterona
ocorre o aparecimento das características sexuais femininas.
103.
102
EXERCÍCIOS
1 – Diferenciereprodução sexuada de assexuada. Qual destas produz maior
variabilidade genética?
2 – Explique por quê o sexo masculino na espécie humana é denominado
de heterogamético.
3 – A análise do cariótipo de certa espécie animal revelou a existência de
50 cromossomos nas células somáticas da fêmea, enquanto o macho
apresentava sempre 49. Que tipo de determinação do sexo ocorre em
tal espécie?
4 – Em drosófila, qual é o sexo esperado de cada uma das seguintes combi-
nações de autossomos (A) e cromossomos X:
a) 3A, XY
b) 2A, 3X
c) 2A, XY
d) 2A, 2X
5 – Na abelha Apis melífera as células somáticas das fêmeas apresentam 32
cromossomos, enquanto as dos machos apresentam 16 cromossomos.
Explique a origem das fêmeas e dos machos com esse número de cro-
mossomos.
6 – Quando o mesmo individuo forma gametas masculinos e femininos este
é chamado de:
a) heterógeno.
b) bipolar.
c) hermafrodita.
d) heterozigoto.
e) dioico.
104.
103
7 – Umaespécie que apresenta indivíduos produtores de gametas femininos
e indivíduos produtores de gametas masculinos é denominada de:
a) bipolar.
b) heterozigota.
c) hermafrodita.
d) dioica.
e) monoica.
8 – Em uma espécie de inseto, as fêmeas possuem 20 cromossomos nas suas
células somáticas. Sabendo-se que nessa espécie o sistema de determi-
nação do sexo é do tipo XX:X0, é esperado que:
a) 100% dos óvulos tenham 10 cromossomos e que 100% dos esperma-
tozoides tenham 9 cromossomos
b) 100% dos óvulos e 100% dos espermatozoides tenham 10 cromos-
somos.
c) 100% dos óvulos e 50% dos espermatozoides tenham 10 cromossomos
e 50% dos espermatozoides tenham 9 cromossomos.
d) 100% dos espermatozoides e 50% dos óvulos tenham 10 cromossomos
e 50% dos óvulos tenham 9 cromossomos.
e) nenhuma alternativa está correta.
9 – A característica mais fundamental da vida é a reprodução, processo
pelo qual os seres vivos têm se perpetuado em nosso planeta desde sua
origem. Com relação aos sistemas de determinação do sexo, assinale a
alternativa falsa:
a) O sistema XY determina o sexo de muitas espécies dioicas, tanto
animais como vegetais.
b) Nos sistemas XY e X0, as fêmeas constituem o sexo heterogamético.
c) No sistema ZW, machos e fêmeas diferem entre si quanto a um par
de cromossomos e são as fêmeas que possuem o par heteromórfico.
d) No sistema X0, os machos têm número ímpar de cromossomos no
cariótipo, apresentando um cromossomo a menos que as fêmeas.
e) No sistema haplodiploide, os machos são portadores de apenas um
lote de cromossomos.
105.
104
10 – Analiseas seguintes afirmativas sobre a determinação do sexo em
abelhas e zangões.
I. A produção de zangões caracteriza o processo de reprodução asse-
xuada denominado de partenogênese.
II. O sistema de determinação do sexo é denominado haplodiploide, já
que as fêmeas possuem dois lotes cromossômicos (2n) e os machos
apenas um (n).
III. Os zangões podem ser considerados clones perfeitos da abelha rainha,
uma vez que não houve a fecundação dos óvulos que os originaram.
Da análise das afirmativas acima podemos assegurar que:
a) apenas I está correta.
b) I e II estão corretas.
c) II e III estão corretas.
d) apenas II está correta.
e) I e III estão corretas.
106.
105
CAPÍTULO VI
ALTERAÇÕES CROMOSSÔMICAS
FernandaCristina Bezerra Leite
Cromossomos: composição e estrutura
Os cromossomos são estruturas celulares compostos por ácidos
nucléicoseproteínasformandoacromatina.Emorganismoseucariotos,
os cromossomos estão armazenados no núcleo celular e, nas células
procariotas, como as bactérias, estes cromossomos estão situados na
região denominada nucleóide, a qual difere do núcleo eucarioto por
não estar delimitada por uma membrana. As proteínas cromossômicas
podem ser do tipo histonas e não-histonas, sendo as proteínas histonas
as de função relacionada à estrutura da cromatina. As histonas são
classificadas em cinco tipos, H1, H2a, H2b, H3 e H4 e, a partir de sua
interação com o filamento de DNA, formam estruturas denominadas
nucleossomos, os quais representam a subunidade cromossômica.
Esta interação histonas-DNA resulta na compactação da informação
genética contida na molécula de DNA, permitindo, por exemplo, que
o genoma humano com aproximadamente de 1,8m de comprimento
(somando todos os filamentos de DNA) possa estar confinado ao
núcleo celular. Esta compactação cromossômica pode se apresentar
organizada em três níveis distintos que irão depender das interações
entre estas proteínas e o estado metabólico da célula, o que determina
se os cromossomos estarão ativos ou inativos.
Cromossomos metafásicos e o cariótipo
Os cromossomos apresentam-se em diversos estados de con-
densação a depender do estágio do ciclo celular em que a célula se
encontra e ainda do nível transcricional dos genes que o compõem,
podendo apresentar-se de forma estrutural mais relaxada, quando há
expressão gênica, ou mais condensada, quando a expressão dos genes
está inativada. A condensação cromossômica também é característica
do estágio metáfase do ciclo celular, uma vez que neste estágio há
107.
106
migração dos cromossomoshomólogos para pólos celulares opostos,
eventoestequeantecedeacitocinese.Estacondensaçãocromossômica
facilitaacorretadistribuiçãocromossômicaentreascélulasfilhase,nesta
fase, os cromossomos encontram-se no maior nível de compactação,
apresentando-se como estruturas cromossômicas bem individualiza-
das, os cromossomos metafásicos. Por este motivo, o grau máximo de
condensação, é que os cromossomos metafásicos são mais atrativos e
indicadosparaestudoscitogenéticos,atravésdosquaisépossívelestudar
o conjunto cromossômico de um indivíduo, o cariótipo, bem como as
alterações cromossômicas que este cariótipo possa apresentar.
Ocariótipoéadescriçãodoconjuntocromossômicodeumadetermi-
nadaespécie,considerando-secaracterísticascomonúmero,tamanhoe
posiçãodocentrômeronoscromossomos.Onúmerobásicodecromos-
somosqueformaocariótipodeumumaespécieédenominadogenoma
haplóide,eesteconjuntohaplóidepodeestarrepresentadomaisdeuma
vez, a depender da programação genética do indivíduo. Tomemos por
exemploaespéciehumana:ascélulasgaméticasapresentamapenasum
conjunto cromossômico (n) formado por 23 cromossomos, no entanto
a maiorias das células apresentam 46 cromossomos e, portanto, são
diplóides (2n), pois apresentam duas cópias de cada cromossomo, ou
seja, um conjunto cromossômico duplicado. Um exemplo de variação
cariotípica programada geneticamente acontece nas células hepáticas,
pois estas células possuem 92 cromossomos, o que resulta em células
tetraplóides (4n). Cada espécie possui em cariótipo específico e o nú-
mero de cromossomos de seus cariótipos não está relacionado nem a
complexidade nem ao tamanho deste organismo. Entretanto, técnicas
citogenéticas podem revelar alterações neste conjunto cromossômico.
Estasalteraçõespodemsergeneticamenteprogramadas,comooexemplo
das células hepáticas, ou não.
Alterações cromossômicas: Numéricas e Estruturais
As alterações cromossômicas não programadas geneticamente,
também chamadas de mutações cromossômicas, podem resultar em
108.
107
mudanças fenotípicas noindivíduo portador. Estas alterações podem
ocorrerquantoaonúmerodecromossomosnumacélulacomotambém
por alterações em determinado fragmento cromossômico.
As alterações que causam mudanças no número cromossômico
(ploidia) de uma célula ou indivíduo são denominadas alterações numé-
ricas. Estas podem ser resultado tanto da adição de conjuntos cromos-
sômicos completos, poliploidia, e resultam em indivíduos portadores de
múltiplos inteiros de conjunto básico cromossômico (haplóide), quanto
da alteração por adição ou perda de uma unidade cromossômica, o que
caracteriza a aneuploidia. As alterações cromossômicas numéricas são
causadas por erros de distribuição cromossômica durante a divisão ce-
lular, por não-disjunção de cromossomos homólogos ou ainda a partir
da fecundação com gametas com número irregular de cromossomos.
As alterações que acontecem em determinados fragmentos de um
cromossomo são denominadas alterações estruturais e resultam de
quebras e rearranjos na estrutura de um mesmo cromossomo como
também entre cromossomos diferentes, sejam eles homólogos ou
não. Estas quebras cromossômicas acontecem por diversos motivos,
dentre eles, desequilíbrio fisiológico ou ainda exposição excessiva a
agentes mutagênicos como radiação ionizante (X e alfa), radiação
ultravioleta e ainda diversos compostos químicos.
Alterações Numéricas
As principais alterações numéricas podem ser agrupadas em duas
categorias: as euploidias e as aneuploidias. Os cariótipos euplóides
apresentam número cromossômico igual a um múltiplo do conjunto
básico e podem ser consideradas normais ou aberrações, a depender
da espécie. Por exemplo, células animais apresentam células em di-
ferentes estados de ploidia como os gametas (haploidia) e as células
hepáticas (poliploidia), sendo a grande maioria das células diplóides.
Nestes casos, estas euploidias são ditas normais, ou geneticamente
programadas. Entretanto, as euploidias não programadas são ditas
aberrantes ou anormais.
109.
108
De acordo como número de conjuntos cromossômicos básicos
apresentados, os cariótipos podem ser sub-classificados em:
a) Monoplóide – organismos que apresentam um único conjunto
básico de cromossomos, sendo representados por (n);
b) Diplóide – cariótipo apresenta dois conjuntos básicos cromos-
sômicos (2n);
c) Triplóide – cariótipo apresenta três conjuntos básicos cromos-
sômicos (3n);
d) Tetraplóide – cariótipo apresenta quatro conjuntos básicos
cromossômicos (4n)
Os indivíduos cujo cariótipo apresenta mais de dois conjuntos
cromossômicos básicos podem ainda serem denominados poliplóides.
Na natureza, pode-se encontrar diversos exemplos de poliploidias
normais, os quais são geneticamente programados para este grau
de ploidia, sendo bastante comum em plantas, porém muito rara em
animais. Em alguns organismos, a poliploidização é tecido-específica
e pode ser resultado da necessidade de cópias extras de determinados
cromossomos e genes que o compõem, como acontece nas células
hepéticas e renais em humanos, por exemplo.
As aneuploidias são caracterizadas pela alteração na quantidade
de apenas um cromossomo, ou parte dele, e não de um conjunto
básico inteiro. Assim, os organismos aneuploides podem apresentar
cromossomos a mais ou a menos em seu cariótipo quando comparado
ao cariótipo do indivíduo normal. Alterações deste tipo resultam da
não disjunção entre cromossomos homólogos ou cromátides durante
as divisões celulares e causam um desbalanço na dosagem gênica,
geralmente resultando em efeito fenotípico. A dose extra cromossô-
mica em determinado indivíduo pode ser denominada de trissomia
cromossômica e é representada por cariótipo2n+1. Na espécie humana,
a trissomia do cromossomo 21 resulta fenotipicamente na síndrome
de Down, a trissomia mais conhecida para a nossa espécie. Várias ou-
110.
109
tras trissomias naespécie humana já foram relatas, porém são letais
e os indivíduos não ultrapassam a fase embrionária pois não toleram
o desbalanço na dosagem cromossômica. Nos casos em que ocorre
a ausência de um dos cromossomos quando comparado ao conjunto
básico cromossômico, esta aneuploidia é denominada monossomia e
representada por 2n-1.
Alterações Estruturais
Uma segunda classe de alterações cromossômicas são as estrutu-
rais que causam um rearranjo no cariótipo do indivíduo e resultam de
quebras na dupla-fita da molécula de DNA sendo seguido da ligação
destes fragmentos, gerando o novo perfil de arranjo cromossômico.
As alterações estruturais podem ocorrer por meio natural, através
do crossing-over, ou por indução por agentes ionizantes como raios
X e gama, por exemplo. O crossing-over é um tipo de recombinação
homóloga que ocorre naturalmente nos organismos e resulta na troca
de fragmentos entre cromossomos.
As alterações cromossômicas estruturais podem ter efeito ba-
lanceado ou desbalanceado, a depender do conteúdo de informação
genética final que resulta destes rearranjos. Por exemplo, se a quebra
cromossômicaresultaemperdaouacréscimodedeterminadofragmen-
to cromossômico, isto irá resultar num desbalanço da dosagem gênica
para este indivíduo. Por outro lado, se a quebra entre cromossomos
homólogos e seguida de troca de fragmento entre estes cromosso-
mos, não haverá perda de informação genética e, conseqüentemente,
não haverá desbalanço gênico causado pelo novo arranjo estrutural.
Assim, as alterações cromossômicas desbalanceadas podem ser
do tipo deleção e duplicação, enquanto as alterações desbalanceadas
podem ser classificadas em inversões, translocações e transposições.
a) Deleção
A deleção é caracterizada pela perda de determinado fragmento
cromossômico e exige que duas quebras cromossômicas aconteçam
111.
110
ladeando este fragmento.A perda da informação genética referente
a este fragmento dá-se pelo fato do mesmo não possuir centrô-
mero, o que impossibilita a correta migração e distribuição deste
fragmento durante o próximo ciclo celular, assim este fragmento
será perdido. O efeito da deleção de um fragmento cromossômico
dependerá do tamanho deste fragmento bem como de sua com-
posição gênica. Deleções que ocorrem em regiões intergênicas são
dificilmente percebidas enquanto as deleções que englobam muitos
genes (multigênicas) têm efeitos muito mais graves que aquelas
intragênica, caracterizada pela perda de um fragmento pertencente
a um único gene.
b) Duplicação
É caracterizada pela repetição de determinado segmento de um
cromossomo. Estas repetições podem estar localizadas adjacentes
entre si ou dispersa em outro local genômico, são as duplicações em
tandem ou insercional, respectivamente. As repetições em tandem
são bastante comuns nos organismos e tem sido utilizadas como
marcadores moleculares para estudos genéticos. Entretanto, se
determinadas duplicações ocorrem em segmentos gênicos, elas
podem causar desbalanços na dosagem gênica e seus efeitos feno-
típicos podem ser perceptíveis.
c) Inversão
Para que ocorra a inversão, deve obrigatoriamente haver dois
pontos de quebra cromossômica e este fragmento é religado de forma
invertida, ápós sofrer um giro de 180º, o que resulta na inversão da
ordem dos genes para o cromossomo em questão. Por se tratar de
uma alteração cromossômica balanceada, não há perda de informação
genética total, exceto nos casos em que a quebra ocorra em regiões
codificadoras (gênicas). Entretanto, a depender da extensão da região
invertida, seus efeitos poderão ser percebidos durante o pareamento
de cromossomos.
112.
111
Podem ser subdivididas,quanto à composição do fragmento in-
vertido em pericêntrica, quando o fragmento cromossômico invertido
contém o centrômero ou paracêntrica, nos casos em que o fragmento
invertido não contém o centrômero, ou seja, compreende apenas a
região de um braço cromossômico.
b) translocação
A translocação ocorre a partir de quebras cromossômicas em que
o fragmento cromossômico irá se unir a outro cromossomo resultando
na transferência de material genético entre eles, troca intercromos-
sômica. Quando esta troca de fragmentos cromossômicos ocorre
entre cromossomos não homólogos e sem haver perda de informação
genética, esta translocação é denominada translocação recíproca,
pois há troca de partes cromossômica entre eles. Entretanto, se um
fragmento é translocado para outro cromossomo, caracterizando
um cromossomo doador e um receptor, esta translocação é simples
ou não recíproca. Em ambos os casos, e resultado é a distribuição
diferenciada de material genético entre os cromossomos, mas não
há perda de informação genética.
c) Transposição
A transposição envolve um único cromossomo e refere-se a trans-
ferência de um fragmento cromossômico de uma região a outra, dentro
do mesmo cromossomo, ou seja, é intracromossômica. De fato, alguns
autores não consideram esta alteração cromossômica por apresentar-
se de forma muito rara, pois para que esta aconteça, é necessário que
haja três quebras cromossômicas num mesmo cromossomo, duas
delas ladeando o fragmento transposto e outra quebra no ponto de
inserção deste fragmento.
113.
112
EXERCÍCIOS
1 – Diferencieas alterações cromossômicas numéricas e estruturais.
2 – Sobre as alterações cromossômicas, assinale (V) para verdadeiro e (F)
para falso:
( ) Os cromossomos são classificados, de acordo com a posição do
telômero, em metacêntrico, submetacêntrico, acrocêntrico e te-
locêntrico.
( ) As alterações cromossômicas podem ser numéricas ou estruturais.
( ) As alterações numéricas são melhor toleradas pelos organismos,
com raras exceções.
( ) Alterações do tipo inversão, duplicação e deleção não causam perda
da informação genética
3 – O cariótipo é a descrição do conjunto cromossômico de uma determi-
nada espécie. Sobre as alterações que podem ocorrer neste conjunto
cromossômico, assinale (V) para verdadeiro e (F) para falso:
( ) A translocação só pode ocorrer entre dois cromossomos homólogos.
( ) O cariótipo representa as características do conjunto cromossômico
de uma determinada espécie.
( ) Os cromossomos representados no cariótipo podem estar em
qualquer fase do ciclo celular
( ) O ideograma é a representação esquemática de um cariótipo
( ) A aneuploidia é caracterizada pela adição ou perda de um ou poucos
cromossomos
4 – Em que diferem as alterações balanceadas e não-balanceadas? Cite
exemplo para cada uma delas.
114.
113
5 – Diferencietranslocação recíproca da não-recíproca.
6 – Descreva exemplos para as alterações numéricas a seguir:
a) Aneuploida
b) Trissomia
c) Poliploidia geneticamente programada
d) Poliploidia anormal
115.
114
CAPÍTULO VII
ESTRUTURA EREPLICAÇÃO DO DNA
Maria de Mascena Diniz Maia e Hildson Dornelas Angelo da Silva
Estrutura do DNA
No ano de 2003, o mundo científico comemorou 50 anos da des-
coberta da estrutura em dupla hélice do ácido desoxirribonucleico, o
tão famoso ADN (DNA - sua sigla do inglês). Em 1953 os estudiosos
James Watson e Francis Crick, utilizando os conhecimentos obtidos por
Miescher, Kossel, Levene e Chargaff, sobre a constituição química do
DNA, propuseram que a estrutura tridimensional do DNA consistia em
dois filamentos de nucleotídeos unidos antiparalelamente (Figura 1).
Figura 1 – Fonte: http://de.wikipedia.org/wiki/
Desoxyribonukleins%C3%A4ure#mediaviewer/File:DNA_Overview.pn
Fonte da Informação Genética
A descoberta da estrutura do DNA e sua responsabilidade como
detentora das informações genéticas foram resultados de diversas
pesquisas científicas que começaram em meados do século XIX com
o isolamento do ácido desoxirribonucleico por Friedrich Miescher.
Um dos principais passos para a identificação das funções do
DNA foi realizado por Frederick Griffith, que identificou o chamado
princípio transformante ao estudar Streptococcus pneumoniae e o
Chargaff,
sobre
a
constituição
química
do
DNA,
propuseram
que
a
estrutura
tridimensional
do
DNA
consistia
em
dois
filamentos
de
nucleotídeos
unidos
antiparalelamente
(Figura
1).
Fonte
da
Informação
Genética
A
descoberta
da
estrutura
do
DNA
e
sua
responsabilidade
como
detentora
das
informações
genéticas
foram
resultados
de
diversas
pesquisas
científicas
que
começaram
em
meados
do
século
XIX
com
o
isolamento
do
ácido
desoxirribonucleico
por
Friedrich
Miescher.
Um
dos
principais
passos
para
a
identificação
das
funções
do
DNA
foi
realizado
por
Frederick
Griffith,
que
identificou
o
chamado
princípio
transformante
ao
estudar
Streptococcus
pneumoniae
e
o
seu
processo
de
transformação.
Esse
foi
o
primeiro
indício
do
armazenamento
de
informações
no
DNA,
para
tal,
Griffith
investigou
a
passagem
de
informações
entre
bactérias
ao
injetar
bactérias
tipo
IS
vivas
Figura
1.
Fonte:
http://de.wikipedia.org/wiki/Desoxyribonukleins%
C3%A4ure#mediaviewer/File:DNA_Overview.pn
g
116.
115
seu processo detransformação. Esse foi o primeiro indício do arma-
zenamento de informações no DNA, para tal, Griffith investigou a
passagem de informações entre bactérias ao injetar bactérias tipo IS
vivas (causadora de pneumonia) nos camundongos após um período
de tempo, os mesmos morreram, enquanto as bactérias tipo IIR vivas
(não causadoras de pneumonia) não causavam morte nos animais
infectados. Quando as bactérias tipo IS foram mortas por aquecimen-
to e posteriormente injetadas nos camundongos, eles continuavam
vivos. Por outro lado, ao se misturar bactérias vivas do tipo IIR com
bactérias mortas do tipo IS e inoculava no animal, após um período
estes morreram. Com isso, Griffith defendeu que nas bactérias tipo
IS existia um fator transformante que seria responsável por transmitir
informações que causaria a mudanças nas bactérias tipo IIR para IS e
assim a virulência (Figura 2).
Figura 2 – Experimento do princípio transformante de Frederick Griffith.
Figura 2. Experimento do princípio transformante de Frederick Griffit
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
Figura 2. Experimento do princípio transformante de Frederick Griffith.
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
IIR vivas
Camundongo injetado com bactérias IS
Figura 2. Experimento do princípio transformante de Frederick Griffit
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
IIR vivas
Figura 2. Experimento do princípio transformante de Frederick Griffith.
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
IIR vivas
Camundongo injetado com bactérias IIR
Figura 2. Experimento do princípio transformante de Frederick Griffit
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
Figura 2. Experimento do princípio transformante de Frederick Griffith.
Funções do Material Genético
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
IIR vivas
Camundongo injetado com bactérias tipo IS mortas por aquecimentos
Figura 2. Experimento do princípio transformante de Frederick Griffit
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
Figura 2. Experimento do princípio transformante de Frederick Griffith.
Funções do Material Genético
Camundongo injetado com bactérias
IS
Camundongo injetado com bactérias
IIR
Camundongo injetado com bactérias
tipo IS mortas por aquecimentos
Camundongo injetado com bactérias
tipo IS mortas por aquecimento e tipo
IIR vivas
Camundongo injetado com bactérias tipo IS mortas por aquecimento e tipo IIR vivas
117.
116
Esse experimento nãoconcluiu o DNA como detentor das infor-
mações genéticas, mas aponta para o material genético. A compro-
vação deste fato ocorre quando Oswald Avery e seus colaboradores
Colin MacLeod e Maclyn McCarty, utilizam enzimas específicas para
degradar proteínas, RNA e DNA (proteases, rinbonucleases e deso-
xirribonucleases, respectivamente) e observam que a capacidade
transformante só foi inibida ao ser utilizada as desoxirribonucleases.
Posteriormente, com o uso de eletroforese, ultracentrifugação e
espectroscopia de ultravioleta puderam confirmar que o princípio
transformante era o DNA.
Contudo, alguns pesquisadores mais céticos levantaram a hipótese
que uma pequena quantidade de proteína pode ter permanecida ligada
ao DNA e não ter sofrido degradação. Outros pesquisadores como
Alfred Hershey e Martha Chase utilizando bacteriófagos marcados
com enxofre (S35
) e fósforo (P32
) radioativos chegaram à mesma
conclusão de Avery.
Funções do Material Genético
Os ácidos nucléicos são as maiores moléculas encontradas no
mundo vivo. São responsáveis pelo controle dos processos vitais bá-
sicos em todos os seres, e para o desempenho de tais funções, o DNA
precisa apresentar três características essenciais.
Armazenamento complexo de informações. O material genético
deve manter guardada uma enorme quantidade de informações
que serão utilizadas na instrução para todas as caraterísticas e
funções de um ser vivo.
Transmissão fiel da informação. Durante o crescimento de um
organismo, a multiplicação celular é algo que ocorre com com-
plexidade, e a cópia das informações contidas no DNA necessita
ser precisa. Essa fidelidade é à base da replicação do DNA e da
transmissão que acontece a cada duplicação da célula.
118.
117
Controle do desenvolvimento.Parte das informações contidas no
DNA será utilizada na codificação de características (no fenótipo).
As proteínas que são codificadas a partir da tradução das instru-
ções genéticas contidas no material genético serão responsáveis
pelo desenvolvimento e diferenciação celular, características que
possibilitam a diversidade de tipos celulares, mesmo que em um
mesmo organismo.
Estrutura Primária
A estrutura primária de uma molécula de DNA consiste em uma
sequência de nucleotídeos (Figura 3). Os nucleotídeos unem-se por
ligações fosfodiéster e são compostos por três componentes – um
grupo fosfato, uma pentose (desoxirribose no DNA e ribose no RNA)
e uma base nitrogenada.
Componentes dos Nucleotídeos
O grupo fosfato (PO4
). É derivado do ácido fosfórico (H3
PO4
) – é
comum tanto ao DNA como ao RNA. Tem a função de ligar os
nucleotídeos de uma mesma fita e é responsável por produzir uma
carga negativa, tornando o DNA e RNA de caráter ácido.
Pentose. Açúcar que apresenta cinco carbonos enumerados de
1’ a 5’. Quatro átomos de carbono e um de oxigênio são unidos
para formar um anel de cinco lados. No carbono 3’ a presença de
um átomo de hidrogênio (desoxirribose, presente do DNA) ou
uma hidroxila (ribose, presente no RNA) é responsável pela ligeira
11
utilizada na codificação de características (no fenótipo). As proteínas que sã
codificadas a partir da tradução das instruções genéticas contidas no materia
genético serão responsáveis pelo desenvolvimento e diferenciação celula
características que possibilitam a diversidade de tipos celulares, mesmo qu
em um mesmo organismo.
Estrutura Primária
A estrutura primária de uma molécula de DNA consiste em um
sequência de nucleotídeos (Figura 3). Os nucleotídeos unem-se por ligaçõe
fosfodiéster e são compostos por três componentes – um grupo fosfato, um
pentose (desoxirribose no DNA e ribose no RNA) e uma base nitrogenada.
Fosfato
Pentose Base
Nitrogenada
Figura 3. Esquema de um nucleotídeo.
Figura 3. Esquema de um nucleotídeo.
119.
118
diferenciação entre osnucleotídeos do DNA e RNA, porém essa
pequena diferença é identificada por todas as enzimas envolvidas
no metabolismo dos ácidos nucleicos.
Base Nitrogenada. A classificação das bases nitrogenadas se dá
pela presença de um ou dois anéis heterocíclicos. As bases púricas
ou purinas apresentam dois anéis e possuem duas representantes,
adenina (A) e guanina (G), que se encontram tanto no DNA quanto
no RNA. Enquanto as pirimídicas ou pirimidinas que só detêm um
anel, como representantes dispõem da citosina (C) (presente no
DNA e RNA), timina (T) (presente apenas do DNA) e uracila (U)
(presente apenas no RNA) (Figura 4).
DNA é uma hélice
Figura 4. Fonte: http://upload.wikimedia.org/wikipedia/commons/thumb/d/de/RNA-Nucleobases.svg/2000px-RNA-
Nucleobases.svg.png
Figura 4 – Fonte: http://upload.wikimedia.org/wikipedia/commons/thumb/d/de/
RNA-Nucleobases.svg/2000px-RNA-Nucleobases.svg.png
120.
119
Complementariedade das BasesNitrogenadas
A utilização de cromatografia em papel foi realizada por Erwin
Chargaff e seus colaboradores E. Vischer e S.Zamenhof para iden-
tificar a proporção entre as bases nitrogenadas. Os pesquisadores
observaram que em diferentes espécies a quantidade de Adenina era
igual à de Timina e a de Citosina a de Guanina. Essa informação foi de
extrema importância na elucidação da dupla hélice, essa conclusão é
conhecida como a proporção de Chargaff.
DNA é uma hélice
Os experimentos com difração de
raio X inicialmente conduzidos por Mau-
rice Wilkins e aprimorados por Rosalind
Franklin concluíram que o DNA consistia
de uma molécula em hélice (Figura 5). Ao
lado uma imagem do DNA por análise de
difração de raio X.
A “Descoberta” da Dupla Hélice
Com todos os conheci-
mentos sobre o DNA gerado
até então, James Watson e
Francis Crick (Figura 6) fo-
ram capazes de reunir as
informações e elucidar a du-
pla hélice do DNA, que seria
formada por duas cadeias de
polinucleotídeos orientados
antiparalelamente e que in-
teragem por interações de
hidrogênio (pontes de hidro-
gênio). Com giro para a direita e apresenta dois sulcos um maior maior
e um menor. Vale salientar que todas essas conclusões são resultados
115
DNA é uma hélice
Os experimentos com difração de raio X
inicialmente conduzidos por Maurice Wilkins e
aprimorados por Rosalind Franklin concluíram
que o DNA consistia de uma molécula em hélice
(Figura 5). Ao lado uma imagem do DNA por
análise de difração de raio X.
Figura 4. Fonte: http://upload.wikimedia.org/wikipedia/commons/thumb/d/de/RNA-Nucleobases.svg/2000px-RNA-
Nucleobases.svg.png
Figura 5. Fonte:
http://i202.photobucket.com/albums/aa1
44/
Primate_bucket/dna.jpg
Figura 5 – Fonte: http://i202.
photobucket.com/albums/aa144/
Primate_bucket/dna.jpg
la Hélice
nhecimentos sobre o DNA gerado até então, James
k (Figura 6)
reunir as
dupla hélice
ada por duas
nucleotídeos
mente e que
rações de
hidrogênio).
e apresenta
maior e um
que todas
esultados em
união de
endo descobertas por vários pesquisadores.
onal da dupla hélice configura a estrutura secundária
Figura 6. Fonte: http://cdn2.hubspot.net/hub/237126/file-660502209-
jpg/watson_crick.jpg
Figura 6 – Fonte: http://cdn2.hubspot.
net/hub/237126/file-660502209-jpg/
watson_crick.jpg
121.
120
em grande parteda união de informações que foram sendo des-
cobertas por vários pesquisadores.
Estrutura Secundária
A forma tridimensional da dupla hélice configura a estrutura
secundária da molécula de DNA. Os dois filamentos são mantidos
próximos devido a forças moleculares conhecidas como interações
de hidrogênios. A quantidade de interações de hidrogênio entre
os nucleotídeos de fitas complementares depende dos tipos en-
volvidos. Entre guaninas e citosinas são encontradas três intera-
ções, enquanto duas entre adeninas e timinas, por isso ocorre um
pareamento mais forte entre G – C do que entre A – T.
Formas Alternativas da Dupla Hélice
A dupla hélice do DNA apresenta uma variação conformacional
dependendo da concentração de sais e da natureza das moléculas
que interagem com o material genético. A estrutura encontra-
da normalmente nas condições fisiológica é chamada de DNA
B. Contudo outras conformações como DNA A, que apresenta
cerca de 11 pares de nucleotídeos por volta e possui uma torção
para a direita (dextrogira) e do DNA Z com aproximadamente
12 pares por volta e torção para a esquerda (levogira) já são
identificadas in vitro.
Replicação do Dna
Todos os organismos unicelulares ou pluricelulares têm a ca-
pacidade de se reproduzirem para a manutenção da espécie, e a
divisão celular está no início desse processo, permitindo a perpe-
tuação da vida. Para que tudo isso seja garantido, a replicação do
DNA tão necessário a esse processo deve ocorrer de forma precisa.
Na replicação do DNA ocorre a separação das fitas parentais e
a produção de duas novas fitas, tendo as parentais como molde.
Cada nova molécula de DNA contém uma fita parental e uma fita
122.
121
recém-sintetizada, caracterizando areplicação semiconservativa.
O processo de replicação é altamente complexo e apresenta o
envolvimento de várias estruturas, principalmente enzimas que
atuam de forma harmônica para garantir uma duplicação fidedigna.
Replicação Semiconservativa
Inicialmente levantava-se a possibilidade de três tipos de re-
plicação – conservativa, semiconservativa e dispersiva – porém,
com os experimento de Matthew Meselson e Franklin Stahl com
o isótopo do nitrogênio (15N) concluiu-se que a mesma ocorria
de forma semiconservativa. Os pesquisadores analisaram os pesos
moleculares das moléculas de DNA recém-sintetizadas e observa-
ram que as novas cadeias apresentavam pesos correspondentes a
uma replicação semiconservativa (uma fita pesada e uma fita leve),
como mostrado na figura abaixo (Figura 7).
rocessos da Replicação
Figura 7. Fonte: https://djalmasantos.files.wordpress.com/2010/10/01-novo.jpg
Figura 7 – Fonte: https://djalmasantos.files.wordpress.com/2010/10/01-novo.jpg
123.
122
Processos da Replicação
Todoo processo ocorre ordenadamente e quase que simultâneo a
fim de prover uma replicação rápida e fiel. Didaticamente é dividido em
três etapas: início, alongamento e término. Abaixo pode-se observar
uma imagem que apresenta os principais componentes do processo
(Figura 8), vale salientar que a perpetuação das espécie e a evolução
dependem diretamente da duplicação celular e assim a replicação
mostra-se tão importante.
Forquilhas de Replicação
As origens de replicação são regiões conservadas em procariotos
e eucariotos e que determinam o local onde se inicia a duplicação do
DNA. Os procariontes apresentam uma única origem de replicação
(OriC em Escherichia coli), já nos eucariontes a presença de vários
elementos ARS (inglês) ou SRA (sequências de replicação autônomas)
indicam o existência de várias origens. O complexo proteico formado
pela DNA polimerase, helicase, primase, ligase e outros interagem
com as origens e formam a forquilha de replicação. A partir das
118
Processos da Replicação
Todo o processo ocorre ordenadamente e quase que simultâneo a fim
de prover uma replicação rápida e fiel. Didaticamente é dividido em três etapas:
início, alongamento e término. Abaixo pode-se observar uma imagem que
apresenta os principais componentes do processo (Figura 8), vale salientar que
a perpetuação das espécie e a evolução dependem diretamente da duplicação
celular e assim a replicação mostra-se tão importante.
Figura 7. Fonte: https://djalmasantos.files.wordpress.com/2010/10/01-novo.jpg
Figura 8. Fonte: https://biolego.files.wordpress.com/2011/10/450px-dna_replication_pt-svg.png
Figura 8 – Fonte: https://biolego.files.wordpress.com/2011/10/
450px-dna_replication_pt-svg.png
124.
123
foquilhas, os complexosproteicos (bolhas de replicação) partem em
sentidos opostos (bidirecionais) (Figura 9). Cada bolha é responsável
pela polimerização de uma parte do DNA chamada de replicon.
Separação das fitas e giro
AdesnaturaçãodaduplafitaocorreacargodaenzimaDNAhelicase
que rompe as interações de hidrogênio entre as bases complementares
e causa uma torção na cadeia do DNA. As enzimas DNA topoisomera-
se I (DNA girase) geram rompimentos nas ligações fosfodiéster para
diminuir a pressão ocasionada pela torção e assim permite o giro total
das fitas do DNA, deixando-as retilíneas e aptas para replicação.
Estabilização da desnaturação
Durante a quebra e giro da cadeia dupla as moléculas de fitas
simples de DNA ficam expostas e susceptíveis a renaturação, a fim de
proteger essas regiões são adicionadas nas fitas proteínas de ligação
ao DNA unifilamentar (proteínas SSB).
Iniciação da replicação
As polimerases, enzimas que replicam efetivamente o DNA,
não conseguem catalisar a síntese desde o início, elas necessitam
1
das foquilhas, os complexos proteicos (bolhas de replicação) partem e
sentidos opostos (bidirecionais) (Figura 9). Cada bolha é responsável pe
polimerização de uma parte do DNA chamada de replicon.
Separação das fitas e giro
A desnaturação da dupla fita ocorre a cargo da enzima DNA helica
que rompe as interações de hidrogênio entre as bases complementares
causa uma torção na cadeia do DNA. As enzimas DNA topoisomerase I (DN
girase) geram rompimentos nas ligações fosfodiéster para diminuir a press
ocasionada pela torção e assim permite o giro total das fitas do DNA, deixand
as retilíneas e aptas para replicação.
Forquilhas de replicação
Figura 9. Fonte: http://images.slideplayer.com.br/1/334307/slides
/slide_14.jpg
Forquilhas de Replicação
Figura 9 – Fonte: http://images.slideplayer.com.br/1/334307/slides/slide_14.jpg
125.
124
de um pequenofilamento de nucleotídeos, um oligonucleotídeo ini-
ciador, conhecido como primer. O primer é formando por seis a trinta
ribonucleotídeos e produzido a partir de um complexo denominado
primossomo, que apresenta uma RNA polimerase (primase) como
principal constituinte. O papel principal do primer é fornecer uma
hidroxila livre (presente no carbono 3’) necessária para que as poli-
merases iniciem o processo.
Polimerização
Após a produção dos primers, as DNA polimerases apresentam
todos os requisitos para de fato iniciar a replicação. Existem várias
DNA polimerases, nos procariontes destacasse as DNA polimerases
I, II e III, enquanto nos eucariontes encontramos as DNA polimerases
α, β, γ, δ e ε. Vale ressaltar que algumas delas são utilizadas no reparo
do DNA e as responsáveis pela replicação efetivamente são as DNA
polimerases II e III (procariontes) e α e δ (eucariontes). O conjunto das
enzimas – polimerases, helicases, topoisomerases, primases, ligase e
outras – formam o replissomo, unidade que vai adicionando nucleo-
tídeos a fita em formação, sempre no sentido 5’ → 3’.
Fragmento de Okazaki e DNA ligase
A orientação antiparalela da cadeia dupla e a incapacidade das
enzimas em polimerizar no sentido 3’ → 5’ faz com que uma das novas
fitas a serem produzidas ocorra de forma contínua (filamento leading),
pois segue o sentido 5’ → 3’ precisando de apenas um iniciador, en-
quanto a fita 3’ → 5’ vai sendo sintetizada em pedaços descontínuos
(filamento lagging) com cerca de 1.000 a 2.000 nucleotídeos nos
procarionte e 100 a 200 nos eucariontes, denominados fragmentos
de Okazaki. Cada pedaço é formando a partir de um iniciador (primer)
e são alongados no sentido funcional 5’→3 (Figura 10).
Posteriormente os primers que estão interrompendo os fragmen-
tos de Okazaki na fita descontínua serão removidos e substituídos
por nucleotídeos de DNA. Para a finalização da fita polimerização,
126.
125
a enzima DNAligase fará a união dos fragmentos de Okazaki aos
nucleotídeos recém-adicionados através de ligações fosfodiéster. O
filamento contínuo possui apenas um primer que também será subs-
tituído pela DNA polimerase.
121
fragmentos de Okazaki. Cada pedaço é formando a partir de um iniciador
(primer) e são alongados no sentido funcional 5’→3 (Figura 10).
Posteriormente os primers que estão interrompendo os fragmentos de
Okazaki na fita descontínua serão removidos e substituídos por nucleotídeos
de DNA. Para a finalização da fita polimerização, a enzima DNA ligase fará a
união dos fragmentos de Okazaki aos nucleotídeos recém-adicionados através
de ligações fosfodiéster. O filamento contínuo possui apenas um primer que
também será substituído pela DNA polimerase.
Figura 10. Fonte: https://djalmasantos.files.wordpress.com/2010/10/08-ocorri.jpg
Figura 10 – Fonte: https://djalmasantos.files.wordpress.com
/2010/10/08-ocorri.jpg
127.
126
EXERCÍCIOS
1 – Abiotecnologia trabalha predominantemente manipulando material
genético de seres vivos; para a maioria dos organismos, o material ge-
nético é a molécula de DNA. Acerca da estrutura da molécula de DNA,
assinale a opção correta.
a) O DNA é composto por quatro tipos de bases nitrogenadas: adenina,
citosina, timina e uracila.
b) O açúcar contido na molécula de DNA é a ribose.
c) Em uma cadeia de DNA, os nucleotídeos se ligam aos açúcares e
fosfatos por pontes de hidrogênio.
d) As duas fitas que compõem a molécula de DNA são ligadas por pontes
de hidrogênio entre suas bases nitrogenadas.
e) Há complementaridade entre as bases nitrogenadas adenina e timina
e entre guanina e uracila
2 – Por que os primers na replicação de DNA in vivo não podem ser de DNA?
a) Porque são muito curtos e as DNA polimerases só sintetizam frag-
mentos grandes
b) Porque devem ser feitos no sentido 3´-5´ e as DNA polimerases não
podem fazer isso
c) Porque não há extremidade 3´-OH disponível para síntese de DNA
d) Porque o heterohíbrido é mais fácil de ser retirado depois de com-
pletada a fita descontínua
e) Porque só ocorrem na fita descontínua
3 – Na replicação do DNA fita dupla é imprescindível a formação de frag-
mentos de DNA recém replicados numa das fitas (chamados fragmentos
de Okazaki) porque:
128.
127
a) a primaseadiciona múltiplos primers numa das fitas;
b) a formação de uma fita única dos dois lados seria instável na natureza;
c) a DNA polimerase sempre faz uma fita contínua e outra descontínua
na replicação de DNA;
d) as fitas são antiparalelas e as DNA polimerases só sintetizam num
sentido;
e) a forquilha de replicação progride apenas num sentido.
4 – Cinco amostras com ácidos nucléicos foram analisadas quimicamente e
apresentaram os seguintes resultados:
I) 1ª. amostra: ribose
II) 2ª. amostra: timina
III) 3ª. amostra: dupla hélice
IV) 4ª. amostra: uracila
V) 5ª. amostra: 20% de guanina e 30% de citosina
Entre estas amostras, quais se referem a DNA?
a) Apenas I e II.
b) Apenas I e III.
c) Apenas II e III.
e) Apenas II e IV.
e) Apenas II e V.
5 – O DNA e o RNA são constituídos de muitas unidades, os nucleotídeos.
Cada nucleotídeo é constituído por um grupo fosfato, uma pentose e
uma base nitrogenada. A diferença entre DNA e RNA está:
a) na pentose e nas bases nitrogenadas.
b) no fosfato e nas bases nitrogenadas.
c) na pentose e no fosfato.
d) na pentose, nas bases nitrogenadas e no fosfato.
e) apenas nas bases nitrogenadas
129.
128
6 – Numamolécula de ADN, a quantidade de...
A) adenina mais timina é igual à de citosina mais guanina.
B) citosina mais uracila é igual à de timina mais adenina.
C) uracila mais adenina é igual à de citosina mais guanina.
D) guanina mais timina é igual à de citosina mais uracila.
E) adenina mais citosina é igual à de guanina mais timina.
7 – Enumere as palavras segundo as indicações certas:
( ) Filamento leanding
( ) Polimerase
( ) Filamento lagging
( ) Polimerase α
( ) Replicação eucariótica
( ) Replicação procariótica
( ) Pontas nos cromossomos
( ) Telomerase
( ) Telômero
( ) Fragmentos de Okazaki
( ) Endonuclease
1) Cadeias que cresce no mesmo sentido da replicação, tem um cresci-
mento continuo.
2) Enzima que produz a quebra no pareamento de bases fazendo assim
com que as hélices se separem.
3) Curtos fragmentos de DNA produzidos por replicação descontinua
eucariótica.
4) Cadeia que cresce no sentido oposto da forquilha de replicação, tem
um crescimento descontinuo.
5) Enzimas fundamentais para a replicação e reparo do DNA
6) Possuem várias origens de replicação, pois tem u m material genético
muito extenso.
130.
129
7) Possuem somenteuma origem de replicação, pois possuem um ma-
terial genético não tão extenso como os eucariontes
8) Replicação eucariótica
9) Enzima responsável pelo inicio da replicação do DNA.
10) Nome dado a ponta que se forma na replicação eucariótica.
11) Enzima responsável pelo crescimento do telômero, ou seja realiza o
pareamento de bases nas pontas dos cromossomos.
8–Assinaleaalternativaquecontémaspalavrasquecompletamafraseabaixo:
Existem cinco tipos principais de bases nitrogenadas: adenina,
_________, citosina,_________ e uracila. As duas primeiras pos-
suem um duplo anel de átomos de carbono e derivam de uma substância
chamada_________, sendo, por isso, denominadas bases_________.
a) Guanina, timina, purina, púricas.
b) Timina, guanina, pirimidina, púricas.
c) Timina, guanina, pirimidina, púricas.
d) Timina, guanina, púricas, pirimídicas.
e) Guanina, timina, purina, pirimidina.
9 – Griffith e Avery, McLeod e MacCarthy, com uma diferença de quase 20
anos, empregaram o mesmo organismo e avaliaram o mesmo fenômeno
em busca da natureza química da informação genética. Assinala com A
ou G o que foi empregado nos experimentos dos grupos de Avery e de
Griffith, respectivamente.
( ) DNAse
( ) camundongos
( ) inativação por calor
( ) placas de Petri
( ) RNAse
131.
130
10 – Areplicação do DNA fita dupla exige que as duas fitas sejam separadas,
o que implica a necessidade de girar o DNA sobre si mesmo milhões de
vezes durante o processo. Se o DNA for circular, a tensão gerada nele pelo
avanço bidirecional da forquilha de replicação é enorme; além disso, no
fim do processo as duas fitas duplas de DNA ficam presas uma a outra
(catenadas). Que enzimas são responsáveis pelo relaxamento da tensão
gerada na replicação e pela decatenação dos DNAs circulares?
a) toposiomerases
b) helicases
c) girases
d) endonucleases
e) telomerases
11 – Os experimentos de Grifith apontaram para o DNA como molécula
informacional. Eles foram baseados:
a) na análise de colônias bacterianas transformadas;
b) no estudo do curso da infecção de bactérias transformadas em ca-
mundongos;
c) na avaliação da inativação de bacteriófagos pelas radiações UV e gama;
d) no estudo da incorporação de radioisótopos nas fitas recém sinteti-
zadas de DNA;
e) na análise de imagens de difração de raio X obtidas de cristais de DNA.
12 - Sobre o DNA e sua replicação são feitas as seguintes afirmações, dê a
soma das afirmações corretas.
01) A base molecular do processo é uma interação espontânea entre as
bases, formando pares aleatórios.
02) A fidelidade da replicação exige que a nova fita mantenha a mesma
seqüência de bases do filamento modelo.
04) A preservação da seqüência nucleotídica de um gene decorre da
inviabilidade de mutações durante a replicação.
132.
131
08) A ocorrênciade várias forquilhas de replicação aumenta o tempo
necessário para a duplicação da molécula.
16) A replicação semiconservativa produz moléculas-filha geneticamente
idênticas à molécula-mãe original
Soma ( )
13 - Defina “origem de replicação”. Porque cada molécula de DNA em cro-
mossomos eucariotos contém múltiplas origens de replicação?
14 - Muitas origens de replicação foram caracterizadas por conter sequên-
cias ricas em A:T. Esses núcleos ricos em A:T têm algum significado
funcional? Qual?
15 - A Replicação do DNA tem regras e mecanismos básicos que se aplicam
a procariotos e eucariotos. Quais são eles?
16 - Esquematize uma forquilha de replicação e indique qual fita de DNA será
usada como molde para a síntese contínua e para a síntese descontínua
das fitas filhas.
133.
132
CAPÍTULO VIII
TRANSCRIÇÃO ETRADUÇÃO
Paulo Roberto Eleutério de Souza
Transcrição
Transcrição é o processo de formação de moléculas de RNA a par-
tir de uma molécula molde de DNA. Diferentemente do processo de
replicação do DNA, na transcrição apenas uma das moléculas de DNA
participará do processo. Na realidade, apenas parte desta molécula,
ou seja, apenas um segmento específico será transcrito, conhecido
como unidade transcricional.
A molécula de RNA diferencia-se da molécula de DNA em alguns
aspectos, são eles: a existência da base nitrogenada uracila (U) no lugar
da base T do DNA; a pentose é a ribose que substitui a desoxirrobose
do DNA; apresenta-se geralmente em fita simples e é menos estável
que a molécula do DNA.
Geralmente apenas um gene ou, no máximo alguns genes são
transcritos em RNA ao mesmo tempo (transcrição de genes individuais
ou incluir genes contíguos), sendo um processo altamente seletivo,
ou seja, genes individuais só são transcritos quando necessários. Con-
tudo, o tamanho de um gene pode variar muito de organismo para
organismo, e até mesmo genes diferentes que se encontram dentro
de um determinado organismo, podendo abranger desde dezenas de
pares de bases (pb) até milhões delas.
Classes de RNA
Existem três classes principais de RNA, formadas a partir da trans-
crição do DNA, são elas: Existem três principais moléculas de RNA
formadas a partir da molécula de DNA, com funções distintas: O
RNA mensageiro, O RNA transportador e o RNA ribossomal. O RNA
mensageiro (mRNA) é a molécula de RNA, formada por um filamento
simples, responsável pela transmissão da informação genética conti-
134.
133
da no DNApara os ribossomos, onde será traduzida em proteína. O
RNA ribossomal (rRNA) é a molécula de RNA responsável pela síntese
dos ribossomos, quando associado a proteínas ribossomais. E o RNA
transportador (tRNA) é a molécula responsável pelo transporte de
aminoácidos para os ribossomos para serem utilizados na síntese
protéica. O tRNA é o menor RNA da célula e tem o formato de folha
de trevo. Em uma das extremidades livres da sua molécula existe uma
sequencia ACC de bases nitrogenadas, onde ocorre associação com
o aminoácido. Na região oposta do trevo existe uma trinca de bases
conhecida como anticódon, que reconhece o códon do mRNA.
O processo de transcrição requer três componentes importantes
1. uma molécula de DNA que servirá de molde;
2. Os substratos necessários para construir uma nova molécula
de RNA;
3. O aparato de transcrição, consistindo em proteínas necessárias
para catalisar a síntese de RNA.
O molde para a síntese de RNA é a molécula de DNA fita dupla.
O filamento de DNA que será transcrito é o filamento que está no
sentido 3´ para 5´ (chamado filamento molde), o outro filamento não
transcrito é chamado filamento código (sentido 5´para 3´). A molécula
de RNA formada terá polaridade inversa à do filamento molde de DNA
(sentido 5´para 3´), porém tem a mesma polaridade e sequência de
bases que as do filamento código, a não ser pela substituição das bases
T no DNA por U no RNA.
Dentro de uma unidade de transcrição existem três importantes re-
giões:umpromotor,umasequênciacodificantedeRNAeumfinalizador.
O promotor determina o local onde a RNA polimerase se encaixa
no DNA para iniciar a transcrição gênica. O promotor é composto por
sequências específicas de nucleotídeos que auxilia no reconhecimento
e na ligação de proteínas do aparelho de transcrição. Nele ocorre a
135.
134
indicação de qualdas fitas do DNA será transcrita, a direção da trans-
crição e o primeiro nucleotídeo a ser transcrito em RNA (numerado
como +1). Por outro lado, os nucleotídeos posicionados antes do sítio
+1 do início da transcrição recebem número negativos (-). A segunda
região importante é a região codificante do DNA que é a sequência de
nucleotídeos que será transcrita em RNA e a terceira região importante
é o finalizador que é a sequência de nucleotídeos que indica onde a
transcrição será finalizada.
Sequências promotoras
Nos promotores bacterianos são encontradas sequências que
variam muito de promotor para promotor e sequências curtas de
nucleotídeos comuns aos promotores, conhecidas como sequên-
cias consenso, tais como: a sequência consenso -10 ou Pribnow
Box (5´-TATAAT-3´) e a sequência consenso -35 (TTGACA). Alguns
promotores bactrerianos possuem uma terceira sequência consenso,
chamada de elemento antecedente (ricas em nucleotídeos A –T).
Em eucariontes, a região promotora dos genes contém uma ou mais
sequências conservadas. A mais próxima do sítio de início da trans-
crição é a sequência TATA (sequência consenso TATAAAA) situada
entre 25 a 30 pb antecedentes ao ponto de início da transcrição. Uma
segunda sequência conservada dentro do promotor é a sequência
CAAT (GGCCAATCT), situada próximo a região -80 do sítio de início da
transcrição. Ainda são conhecidas duas outras sequência conservadas
em eucariontes, a sequência rica em GC (GGGCGG), e a sequência de
octâmeros (ATTTGCAT).
RNA polimerase
AprincipalenzimaresponsávelpelatranscriçãoéaRNApolimarese.
O RNA é sintetizado a partir de trifosfatos de ribonucleotídeos (rNTP)
que são adicionados simultaneamente ao grupo 3´-OH da molécula
de RNA crescente pela ação da RNA polimerase. A síntese do RNA
diferentemente da síntese do DNA não precisa de primer para iniciar.
136.
135
A ação daRNA polimerase é corroborada pela ação de várias outras
proteínas acessórias que auxiliam no reconhecimento do gene correto
a ser transcrito e no posicionamento correto da RNA polimerase na
região promotora do gene.
Uma vez posicionada, a RNA polimerase separa as duas cadeias
do DNA e apenas uma das moléculas participa na síntese do RNA. A
transcrição ocorre de forma unidirecional.
Em bactérias, existe apenas uma única molécula de RNA polimerase
que catalisa a síntese de todas as classes de RNA bacteriano: RNAm,
RNAr e RNAt. Esta proteína é constituída de cinco subunidades: duas
subunidades alfa (α2
), uma subunidade beta (β), uma subunidade beta
primo (β´) e uma subunidade ômega (ω). Esta última não é essencial
para a transcrição, mas auxilia na estabilidade da enzima.
O fator sigma (Ϭ) é um fator essencial no reconhecimento e ligação
da RNA polimerase no sítio promotor bacteriano. Após a ligação deste
fator ao cerne da RNA polimerase ocorre o início da transcrição na
posição correta. Contudo, este fator não tem função no alongamen-
to da transcrição. Assim, após a iniciação da transcrição, o fator Ϭ é
liberado e ocorre o alongamento da cadeia de RNA.
Aspolimeraseeucarióticassãomultiméricascommaisdeumadúzia
de subuinidades. A maioria das células eucariontes possuem três tipos
de RNA polimerase, que serão responsáveis pela transcrição de grupos
específicos de RNA: a RNA polimerase I, que transcreve rRNAs, exceto
rRNA 5S; RNA polimerase II, transcreve pré-RNAm, snoRNA, alguns
miRNA e alguns snRNAs; e a RNA polimerase III, que transcreve RNAt,
rRNAr 5S, alguns snRNAs e alguns miRNAs. Além disso, dois outro tipos
de RNA polimerases foram descritas em plantas, as RNA polimerases
IV e V. A RNA polimerase IV pequenos RNAs de interferência (siRNA)
no núcleo vegetal e possui papel na metilação DNA e na estrutura da
cromatina e a RNA polimerase V, transcreve alguns siRNA e transcritos
não codificadores (antisenso) de genes alvo de siRNA.
Em eucariontes, o reconhecimento do promotor se dá através de
proteínas acessórias que se ligam ao promotor, auxiliando no reconhe-
137.
136
cimento do promotorpela RNA polimerase. Estas proteínas acessórias
são chamadas de fatores de transcrição que formam o aparato basal
da transcrição. Outra classe de proteínas acessórias são chamadas de
proteínas ativadoras transcricionais, que ao se ligarem em sequências
específicas de DNA, resultam em níveis aumentados de transcrição.
Etapas da transcrição
Iniciação – é a etapa referente a montagem do aparato da trans-
crição no promotor do gene e o início da síntese de RNA. Nesta
etapa ocorre reconhecimento do promotor, formação da bolha
de transcrição e ligação do primeiro nucleotídeo a molécula de
RNA em formação;
Alongamento – etapa seguinte ao início da transcrição, quando
ocorreadiçãoapartirdosegundonucleotídeoacadeiaemformação
com deselicoidação do DNA. Nesta etapa a RNA polimerase sofre
uma mudança na conformação e não é mais capaz de se ligar a
sequências consenso do promotor. Em bactérias ocorre liberação
da subunidade sigma (Ϭ) após a iniciação;
Término – finalização do processo da transcrição, com a separação
da molécula de RNA recém sintetisada do DNA molde. Em bacté-
rias, existem dois tipos principais de finalizadores. Os finalizadores
dependentes de rô, são capazes de causar o término da transcrição
apenas na presença de uma proteína auxliar chamada de fator rô.
Os finalizadores independentes de rô, capazes de causar o fim da
transcrição na ausência de rô.
Iniciação da transcrição
Em procariontes, como mencionado anteriormente a subunidade
sigmadaRNApolimeraseéfundamentalparaoreconhecimentoeligação
da RNA polimerase no sítio promotor. Inicialmente, a subunidade sigma
reconhece e se liga a sequência consenso -35. A sequência -10 auxilia o
138.
137
desenrolamentolocalizadonoDNAparaocorreraoprocessotranscricio-
nal.Emeucariontes,éessencialaatuaçãodeváriosfatorestranscricionais
basaisnaregiãopromotoradogeneparainiciaratranscrição.Alémdestes,
outras sequências reguladorasdenominadas de acentuadoras e silencia-
doras modulam a iniciação da transcrição.
Existemváriosfatoresdetranscriçãoqueatuamnaregiãopromotora
de genes eucariontes, são eles: TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH e
TFIIJ. O TFIID reconhece a região consenso TATA, através da sua proteí-
na de ligação ao TATA (TBP) e de várias proteínas associadas à TBP. Em
seguida,TFIIAuni-seaocomplexo,seguidaporTFIIB.ATFIIFassocia-sea
RNApolimeraseIIeemseguidaunem-seaocomplexoTFIID-TFIIA-TFIIB.
ATFIIFcontémumasubunidadequecatalisaodesenrolamentodoDNA.
Posteriomente,aTFIIEuni-seaocomplexodeiniciação,ligando-seaoDNA
na região 3´em relação ao início da transcrição. Por último, dois outros
fatores, TFIIH e TFIIJ, unem-se ao complexo. O TFIIH possui atividade de
helicaseeseguecomaRNApolimeraseIIduranteoalongamento,desen-
rolando os filamentos na região da transcrição (a bolha de transcrição).
Alongamento da Transcrição
O alongamento de cadeias de RNA em procariontes ocorre dentro
da bolha de transcrição (segmento desenrolado de DNA) catalisada
pelo cerne da RNA polimerase, logo após a liberação da subunidade
Ϭ. A RNA polimerase segue desenrolando a dupla fita de DNA após
o sítio de polimerização e reenrola os filamentos complementares de
DNA atrás do sito de polimerização durante o avanço no processo de
transcrição. Em Escherichia coli (E. coli) aproximadamente 18 pares
de nucleotídeos fazem parte de uma bolha de transcrição. A liga-
ção do DNA e da cadeia de RNA em crescimento â RNA polimerase
promove estabilidade ao complexo de transcrição. Em eucariontes,
o alongamento da transcrição se dá após a formação do complexo
inicial da transcrição, quando a RNA polimerase se situa na região
promotora e coloca o primeiro nucleotídeo na fita de RNA nascente.
Os fatores de transcrição auxiliam no desenrolamento das fitas de
139.
138
DNA e aRNA polimerase segue adicionando novos nucleotídeos por
complementariedade de bases.
Término da Cadeia de RNA
Em procariontes, o término da síntese de moléculas de RNA
ocorre quando a RNA polimerase se depara com um sítio de térmi-
no. Neste momento, o complexo de iniciação se dissocia, liberando
moléculas de RNA nascente. Nestas células, existem dois tipos de
sítios término, um dependende da proteína rô e um independente
da proteína rô. Os sítios independentes da proteína rô contém uma
região rica em GC seguida por seis ou mais pares de bases AT, com
a presença de A no filamento molde. Uma questão interessante a
ser colocada é a presença de sequências invertidas na região rica
em GC, pois estas quando transcritas produzem sequências de RNA
unifilamentares que podem formar bases e estruturas em grampo.
Estas estruturas formadas retardam o movimento das moléculas de
RNA polimerase promovendo o término da transcrição. De forma
semelhante, o mecanismo de término nas células dependentes de
rô ocorre a formação de estruturas em forma de grampo e a ter-
minalização da transcrição.
Processamento do RNA
Durante a produção do pré-mRNA, o mesmo sofrerá um conjunto
de modificações conhecidas como processamento do RNA, deixan-
do-o mais protegido para ser encaminhado ao citoplasma, além de
sofre modificações na sequencia do RNA recém formado (edição
e splicing). Abaixo as quatro principais etapas do processamento:
Capeamento do RNA mensageiro
Com o objetivo de proteger o RNAm contra exonucleases pre-
sentes no citoplasma a primeira etapa é o adicionamento de um
revestimento (CAP) ao resíduo terminal 5’ do RNA. Outra função do
CAP é interagir com fatores envolvidos na iniciação da transcrição.
140.
139
O CAP consisteem uma molécula de 7-metilguanosina (7-MG) uni-
da por uma ligação 5´-trifosfato e sua adição normalmente ocorre
quando a cadeia está com cerca de 30 nucleotídeos de tamanho.
Poliadenilação do RNA mensageiro
Para permitir o aumento de tempo de vida do RNA recém-
formada no citoplasma a enzima poli(A)-polimerase adiciona
cerca 200 adeninas na extremidade 3’ do RNAm em formação.
Além de estabilizar o RNAm a cauda poli A também desempenha
um papel no transporte do núcleo para o citoplasma.
Edição do RNA
Outra alteração pós-transcricional é a edição do RNA (RNA
editing, no inglês). O processo de edição consiste em dois modos
de modificação, mudando as estruturas das bases ou adicio-
nando ou removendo monofosfatos de uridina. As mudanças
das bases normalmente se dão pela conversão de citosinas para
uracilas através de uma proteína de ligação ao RNA que remove
grupamento amino das citosinas. Enquanto a inserção ou de-
leção de uracilas (U), que ocorre principalmente em protozoá-
rios, compreende a participação de um RNA guia que serve de
molde e também cede U a fita em formação. Essas alterações
provocam, muitas vezes, a alteração da fase de leitura do RNA,
de forma que apenas os mRNA que passarem por esse editing
serão lidos corretamente.
Splicing
Ao processamento do pré-mRNA em que sua sequência é
modificada, seja pela remoção ou inserção de segmentos de
RNA, denominamos de “splicing”. O pré-mRNA é formado por
sequencias não codificantes (íntrons) e codificantes (éxons), os
íntrons necessitam ser removidos durante o processamento do
RNA, já que não apresentam informações que serão traduzidas
141.
140
em proteínas. Aremoção dos íntrons pode ocorrer por maneiras
distintas, podendo acontecer de forma autocatalítica, na qual o
próprio RNA remove os íntrons ou com o auxílio de um conjunto
de proteína, que constituem o spliceossomo.
O processo de splicing é responsável pela possibilidade de um
pré-RNAm poder gerar diversas proteínas diferentes, destaca-se as
observações em várias espécies a grande quantidade de proteínas
em relação ao número de genes.
Tradução
A tradução é um processo de formação de moléculas de proteínas
a partir de sequências de nucleotídeos existentes em moléculas de
mRNA, seguindo as especificações do código genético. A maquinaria
da síntese proteica envolve a participação de várias macromoléculas:
RNAm, sRNAr, RNAt, polipeptídeos, enzimas ativadoras de aminoá-
cidos e proteínas solúveis que participam da iniciação, alongamento
e finalização da cadeia polipeptídica.
Durante a síntese proteica, que ocorre nos ribossomo há participa-
ção da molécula de mRNA transcrita a partir do DNA; 3 a 5 moléculas
de rRNA (formação dos ribossomos); e 40 a 60 moléculas de tRNA
(carreadores de aminoácidos). A ordem desta sequência de bases do
mRNA especifica a ordem exata da sequência de aminoácidos da pro-
teína formada. Cada aminoácido é transportado para os ribossomos
por uma molécula de tRNA específica, que possuem uma trinca de
bases complementar ao códon do RNAm, chamada de anticódon. As
moléculas de tRNA contém uma sequência CCA na ponta 3´, e o grupo
carboxila (COO) do aminoácido é ligado ao nucleotídeo adenina na
ponta do tRNA. As enzimas aminoacil-tRNA sintetases determinam
o reconhecimento do tRNA pelo aminoácido específico. Existe uma
enzima aminoacil-tRNA sintetase para cada aminoácido que utiliza
ATP como fonte de energia.
142.
141
A constituição dosribossomos é de aproximadamente 50% de
rRNA e 50% de proteínas. Em procariontes, uma molécula de ribos-
somo possui 70S (Svedberg) e é composta por duas subunidades uma
maior de 50S e uma menor de 30S. A subunidade maior é composta
por moléculas de rRNA 5S e 23S associada a 31 proteínas ribossômi-
cas e a subunidade menor é composta por moléculas de rRNA 16S
associada a 21 proteínas ribossômicas. Em eucariontes, a molécula de
ribossomo possui 80S, composto por duas subunidades, uma maior
de 60S e uma menor de 40S. A subunidade maior contém rRNAs 5S,
5,8S e 28S e 49 proteínas ribossômicas e a subunidade menor é com-
posta por 33 proteínas ribossômicas e moléculas de rRNA 18S. Cada
ribossomo contém 3 sítios de ligação a molécula de tRNA: o sítio A
(ou aminoacil) no qual o primeiro aminoácido é adicionado pelo ami-
noacil-tRNA trazido. O sítio P (ou peptidil) recebe o aminoacil-tRNA
ligado ao polipeptídeo em crescimento. E o sítio E (ou de saída) que
liga-se ao tRNA que está saindo sem carga elétrica. Os aminoácidos se
ligam-se ao tRNA através da ativação pela enzima acil-tRNA sintetase.
Existe uma acil-tRNA sintetase para cada aminoácido.
Existem na natureza apenas 20 tipos diferentes de aminoácidos. O
código genético é constituído por 64 códons. Cada códon é composto
por uma trinca de bases presentes no mRNA. Como explicar então
a existência de apenas 20 aminoácidos na natureza se cada códon
especifica um aminoácido?
Na realidade, o código genético possui algumas caraterísticas:
1. O código genético é triplo, cada aminoácido é especificado
por uma sequência de três nucleotídeos;
2. Existem casos em que um mesmo aminoácido é especificado
por até seis códons diferentes. Desta forma, ocorre uma osci-
lação no código genético, sendo este conhecido como redun-
dante ou degenerado. Por exemplo, a Leucina é codificada por
seis códons diferentes (UUA, UUG, CUU, CUC, CUA e CUG),
destes quatro iniciam com CU e dois com UU.
143.
142
3. Ocódigogenéticoéqueeledeterminaoiníciodasínteseproteica,
geralmente determinadopor uma trinca de bases, chamada de
códon de iniciação “AUG” que especifica o aminoácido metio-
nina, salientando-se que em raras ocasiões os códons GUG e
UUG são também utilizados;
4. A partir de um destes códons acima descritos, a sequência
de nucleotídeos é traduzida de forma contínua por grupos de
três nucleotídeos seguindo a matriz de leitura do mRNA, ou
seja, o código genético é geralmente não superposto, sendo
que 03 códons (UAA, UAG e UGA) não especificam nenhum
aminoácido, os códons de terminação da síntese da cadeia po-
lipeptídica. Porém, alguns genes superpostos são encontrados
em vírus, mas os códons que fazem parte de um mesmo gene
não se superpõem;
5. Além disso, o código genético é quase universal, existem al-
gumas exceções. Por exemplo, na mitocôndria de humanos
o códon UGA (especifica triptofano ao invés de um códon de
terminação), o códon AUA (especifica metionina ao invés de
isoleucina), AGA e AGG (especificam códons de terminação ao
invésdearginina).NoDNAbacteriano,ocódonUGAespecifica
triptofano ao invés de códon de terminação e em tripanosso-
mos o códon UGA especifica triptofano ao invés de códon de
terminação em humanos.
Etapas da Síntese Protéica
Inicio
Em procariontes, como Escherichia coli (E. coli), o início da síntese
protéica,ocorreatravésdacombinaçãodeváriasmoléculascomo:tRNA
demetionina(tRNA-met),proteínasdeiniciaçãodatraduçãoIF-1,IF-2e
IF-3,aporçãomenordosribossomos(30S)eumamoléculadetrifosfato
deguanosina(GTP)quejuntasdeslizamsobreomRNAmensageiro,até
encontrar o primeiro códon a ser traduzido, o códon AUG.
144.
143
Primeiramente o mRNAliga-se a subunidade menor dos ribosso-
mos. Uma vez que o fator de iniciação IF3 liga-se a subunidade menor
e impede que a subunidade maior se ligue durante a iniciação. Em se-
guida, o tRNA-met associado ao fator IF-2 e ao GTP atingem o mRNA
ligado a subunidade menor dos ribossomos e deslizam até encontrar o
códon AUG. Esta etapa é conhecida como complexo de iniciação 30S.
Posteriormente, a subunidade maior dos ribossomos (50S) se
liga a subunidade menor (30S) e forma o ribossomo completo 70S.
Quando a subunidade maior se junta ao complexo da inicação, este
passa a ser conhecido como complexo de iniciação 70S. Como exis-
tem vários códons AUG numa sequência de mRNA formada, como
é determinado o códon exato do início da tradução? Existe uma
sequência consenso em procariontes que determina qual o códon
AUG de início da tradução, a sequência Shine-Dalgarno (AGGAGG)
que ocorre a aproximadamente sete nucleotídeos upstream ao códon
de iniciação AUG.
Em eucariontes, o processo de iniciação é semelhante, porém
existem algumas modificações. Não existe a sequência Shine-Dalgarno
no mRNA de em eucariontes. A subunidade menor dos ribossomos
com auxílio de fatores de iniciação reconhecem a estrutura cap na
ponta 5´ do mRNA e aí se liga, a subunidade menor então se move ao
longo do mRNA até atingir o primeiro códon AUG. Porém, para auxiliar
no encontro do códon correto existe a presença de uma sequência
consenso chamada de sequência Kozak que contém o códon de início:
5´-ACCAUGG-3´. É importante observar que em eucariontes existe
um maior número de fatores de iniciação que atuam na separação
das subunidades ribossômicas, reconhecimento do cap 5´no mRNA,
possuem atividades de RNA helicase e outros participam auxiliando o
transporte do tRNA iniciador e metionina ao complexo de iniciação.
Alongamento
Nos ribossomos existem três sítios, o peptidil ou sítio P (polime-
rização da cadeia), o aminoacil ou sitio A (adição de aminoácidos) e
145.
144
a saída ousítio E. O tRNA-met fica ligado ao sítio P do ribossomo e o
sítio A fica desocupado.
A partir de então, inicia-se o processo de alongamento da cadeia
de aminoácidos que ocorre em três etapas. Inicialmente o próximo
tRNA carreando o segundo aminoácido é trazido conforme especifi-
cado pela sequência do códon presente no mRNA e este é ligado ao
Sítio A. Esta ligação é dependente do fator de alongamento Tu (EF-
Tu) ligado ao GTP. Em seguida, ocorre clivagem de GTP em GDP, e o
complexo EF-Tu-GDP é liberado. O fator Ts de alongamento (EF-Ts)
por sua vez promove o regeneramento do EF-Tu-GDP em EF-Tu-GTP,
este processo ocorre de forma semelhante em eucariontes de forma
que o tRNA é carreado até o sítio A.
Na segunda etapa do processo de alongamento ocorre uma ligação
peptídica entre os aminoácidos adjacentes, ligados aos sítios P e A,
catalisado pela enzima peptidil transferase. Agora o tRNA que antes
ocupava o sítio P passa para o sítio E, e é liberado para o citoplasma,
podendo ser recarregado com outro aminoácido. Na terceira etapa,
ocorre o processo conhecido como translocação, no qual os ribossomos
deslocam-se sobre o mRNA e os dois aminoácidos unidos passam a
ocupar o sítio P, deixando o sítio A vazio disponível para entrada de
um novo aminoácido, processo catalisado pela enzima translocase e
requer o fator G do alongamento (EF-G) e a hidrólise de GTP em GDP.
Então, outro tRNA, com um terceiro aminoácido que seja reconhecido
pelo terceiro códon do mRNA, entra no sítio A e ocorre a formação
de outra ligação peptídica entre o segundo e o terceiro aminoácidos.
O tRNA do segundo aminoácido é liberado e o ribossomo desloca-se
até o próximo códon. A cadeia formada por três aminoácidos passa a
ocupar o sítio P, deixando novamente o sítio A vazio.
Término
Este processo de alongamento (tRNA carregado com seu aminoá-
cido ocupam o sítio A, ocorre a formação da ligação peptídica entre
os aminoácidos nos sítios A e P, e os ribossomos deslocam-se para o
146.
145
próximo códon) érepetido diversas vezes até que na molécula de
mRNA seja encontrado um dos códons de término da síntese protéica,
UAA, UAG, ou UGA. Neste momento, nenhum tRNA entra no sítio A
do ribossomo e proteínas chamadas de fatores de liberação ligam-se
ao ribossomo. Existem três fatores de liberação em E. coli: RF1
, RF2
e RF3
. O RF1
liga-se aos códons de término UAA e UAG, e RF2
a UGA
e UAA. O RF3
forma um complexo com GTP e liga-se ao ribossomo.
Em seguida, estes fatores promovem clivagem do tRNA no sítio P
havendo clivagem do GTP em GDP. Fatores adicionais auxiliam na
liberação do tRNA do sítio P, liberação do mRNA do ribossomo e a
dissociação do ribossomo.
Em eucariontes, existem dois fatores de liberação:eRF1
, responsável
pelo reconhecimento de todos os códons de término, e eRF2
que liga
GTP e estimula a liberação do polipeptídeo do ribossomo.
147.
146
EXERCÍCIOS
1 - Durantemuito tempo, os cientistas acreditaram que variações anatô-
micas entre os animais fossem consequência de diferenças significa-
tivas entre seus genomas. Porém, os projetos de sequenciamento de
genoma revelaram o contrário. Hoje, sabe-se que 99% do genoma de
um camundongo é igual ao do homem, apesar das notáveis diferenças
entre eles. Sabe-se também que os genes ocupam apenas cerca de
1,5% do DNA e que menos de 10% dos genes codificam proteínas que
atuam na construção e na definição das formas do corpo. O restante,
possivelmente, constitui DNA não codificante. Como explicar, então, as
diferenças fenotípicas entre as diversas espécies animais?
A resposta pode estar na região não-codificante
do DNA. S. B. Carroll et al. O jogo da evolução.
In: Scientific American Brasil, jun./2008 (com
adaptações).
a) região não-codificante do DNA pode ser responsável pelas diferenças
marcantes no fenótipo porque contém A as seqüências de DNA que
codificam proteínas responsáveis pela definição das formas do corpo.
b) uma enzima que sintetiza proteínas a partir da seqüência de aminoá-
cidos que formam o gene.
c) centenas de aminoácidos que compõem a maioria de nossas proteínas.
d) informações que, apesar de não serem traduzidas em seqüências de
proteínas, interferem no fenótipo.
e) os genes associados à formação de estruturas similares às de outras
espécies.
2 - UFRJ - Suponha um gene de um eucarioto responsável pela síntese de
uma proteína. Nesse gene existem íntrons, ou seja, regiões do ADN cujas
informações não estão presentes na proteína em questão. As regiões
do ARN transcrito correspondentes aos íntrons são eliminadas após o
processo de transcrição. A figura a seguir representa o resultado de
148.
147
uma experiência dehibridação do ARN mensageiro com a cadeia de
ADN que lhe deu origem.
A figura mostra cinco regiões, identificadas por números de 1 a 5.
Quais dessas regiões correspondem aos íntrons?
Justifique sua resposta.
3 - (PUC - SP-2007) A mesma molécula — o RNA — que faturou o Nobel de
Medicina ou Fisiologia na segunda-feira foi a protagonista do prêmio de
Química entregue ontem. O americano Roger Kornberg, da Universidade
Stanford, foi laureado por registrar em imagens o momento em que a
informação genética contida no DNA no núcleo da célula é traduzida para
ser enviada para fora pelo RNA — o astro da semana. Esse mecanismo de
transcrição, através do qual o RNA carrega consigo as instruções para a
produção de proteínas (e por isso ele ganha o nome de RNA mensageiro),
já era conhecido pelos cientistas desde a década de 50.
(Girardi, G. Estudo de RNA rende o segundo Nobel
— O Estado de S. Paulo, 5 out. 2006).
A partir da leitura do trecho acima e de seu conhecimento de biologia
molecular, assinale a alternativa incorreta.
a)AproduçãodeRNAmensageirosedáporcontroledomaterialgenético.
b) No núcleo da célula ocorre transcrição do código da molécula de
DNA para a de RNA.
c) O RNA mensageiro leva do núcleo para o citoplasma instruções trans-
critas a ele pelo DNA.
processo de transcrição. A figura a seguir representa o resultado de uma
experiência de hibridação do ARN mensageiro com a cadeia de ADN que
lhe deu origem.
A figura mostra cinco regiões, identificadas por números de 1 a 5.
Quais dessas regiões correspondem aos íntrons?
Justifique sua resposta.
3 - (PUC - SP-2007) A mesma molécula — o RNA — que faturou o Nobel de
Medicina ou Fisiologia na segunda-feira foi a protagonista do prêmio de
Química entregue ontem. O americano Roger Kornberg, da Universidade
Stanford, foi laureado por registrar em imagens o momento em que a
informação genética contida no DNA no núcleo da célula é traduzida para
ser enviada para fora pelo RNA — o astro da semana. Esse mecanismo de
transcrição, através do qual o RNA carrega consigo as instruções para a
produção de proteínas (e por isso ele ganha o nome de RNA mensageiro),
já era conhecido pelos cientistas desde a década de 50.
(Girardi, G. Estudo de RNA rende o segundo Nobel — O Estado de S.
Paulo, 5 out. 2006).
A partir da leitura do trecho acima e de seu conhecimento de biologia
molecular, assinale a alternativa incorreta.
a) A produção de RNA mensageiro se dá por controle do material genético.
149.
148
d) No citoplasma,o RNA mensageiro determina a sequência de aminoá-
cidos apresentada por uma proteína.
e) Cada molécula de RNA mensageiro é uma longa sequência de nucleo-
tídeos idêntica ao DNA.
4 - O que são sequências intercalares ou íntrons?
5 - O que significa recomposição (splicing) do RNAm?
6 - Conceitue:
a. Codon
b. Anticodon
c. Aminoacil-tRNA sintetase
d. Peptidil transferase
e. Translocase
7 - Quais as características do código genético?
8 - Como se dá o início do processo de tradução
9 - Como se dá o alongamento da tradução?
10 - Como se dá o término da tradução?
11 - Imagine uma molécula de DNA com a seguinte composição de bases:
5´-ATGGGAGCGGATATGCCGGGCAGATAG-3´
a. Qual a sequência complementar desta molécula?
b. Qual a sequência de mRNA formada?
c. Qual a sequência da proteína formada?
150.
149
CAPÍTULO IX
MUTAÇÃO EREPARO DO DNA
Martín Alejandro Montes e Ana Cristina Lauer Garcia
Ao imaginarmos o DNA pensamos muitas vezes em uma molé-
cula estável e de replicação extremamente eficiente. No entanto, um
olhar mais atento nos mostrará que, por vezes, algumas sequências
de nucleotídeos desta incrível molécula da vida podem ser alteradas
de forma natural ou não, processo que chamamos de mutação gênica.
No presente capítulo trataremos da importância das mutações, suas
consequências dependo do local onde ocorrem e dos mecanismos de
reparo que existem para consertar esses erros na molécula de DNA.
Mutações Gênicas: Definição e Importância
Mutações gênicas são modificações nos nucleotídeos de DNA (A,
T, C e G) de um organismo. É o processo pelo qual os genes mudam de
uma forma alélica para outra, portanto, as mutações são consideradas
fonte de variabilidade genética. Embora a maioria das mudanças no
DNA leve a efeitos ruins, a mutação gênica é vital para a evolução. Caso
não ocorresse mutação, o material genético primordial ainda estaria
igual e, possivelmente, ainda seriamos bactérias primitivas. A mutação
fornece novas combinações genéticas, sendo a matéria prima para a
evolução. É sobre a variabilidade gerada pelas mutações e pela recom-
binação que opera a seleção natural, permitindo que as populações
evoluam e se adaptem aos mais diversos tipos de ambientes. Embora
as mutações sejam muito importantes para a adaptação e evolução das
populações e espécies, devemos ter em mente que estas não ocorrem
para beneficio de um organismo em uma determinada situação. As
mutações simplesmente ocorrem e são filtradas pela seleção natural,
de modo que permanecem apenas as adaptativas.
Como Surgem as Mutações?
Dependendo da sua origem, as mutações gênicas podem ser
151.
150
classificadas como espontâneasou induzidas. Mutações espontâneas
são aquelas que ocorrem naturalmente, sem uma causa aparente. As
mutações induzidas são aquelas que aparecem em decorrência da
utilização de agentes mutagênicos.
Entre as mutações espontâneas podemos citar a depurinação e
a desaminação. No primeiro caso uma base púrica (A ou G) de um
nucleotídeo é perdida, gerando um sitio apurínico (Figura 1). Assim,
durante a replicação do DNA, esse sítio não pode atuar como molde
na formação da fita complementar. Como consequência, um nucleo-
tídeo qualquer é incorporado como complementar ao sítio apurínico,
geralmente ocorrendo a incorporação de uma adenina. A desaminação
consiste na perda de um grupo amino (NH2) de uma base nitrogenada.
Essa modificação química altera o padrão de pareamento, por exemplo,
a desaminação da citosina faz com que ela se comporte como uma
uracila e forme par com a adenina durante a replicação.
Uma mutação pode ser induzida por análogos de bases. Estas são
substâncias que tem uma estrutura química similar a uma das bases
nitrogenadas do DNA. Devido a tal similaridade, a DNA polimerase
não consegue distinguir entre a base correta do DNA e o análogo,
podendo incorporar erroneamente este último durante a replicação
aquelas que ocorrem naturalmente, sem uma causa aparente. As mutaçõ
induzidas são aquelas que aparecem em decorrência da utilização de agen
mutagênicos.
Entre as mutações espontâneas podemos citar a depurinação e
desaminação. No primeiro caso uma base púrica (A ou G) de um nucleotíd
é perdida, gerando um sitio apurínico (Figura 1). Assim, durante a replicação
DNA, esse sítio não pode atuar como molde na formação da fita complement
Como consequência, um nucleotídeo qualquer é incorporado co
complementar ao sítio apurínico, geralmente ocorrendo a incorporação de u
adenina. A desaminação consiste na perda de um grupo amino (NH2) de u
base nitrogenada. Essa modificação química altera o padrão de pareamen
por exemplo, a desaminação da citosina faz com que ela se comporte co
uma uracila e forme par com a adenina durante a replicação.
Uma mutação pode ser induzida por análogos de bases. Estas s
substâncias que tem uma estrutura química similar a uma das bas
nitrogenadas do DNA. Devido a tal similaridade, a DNA polimerase n
consegue distinguir entre a base correta do DNA e o análogo, poden
incorporar erroneamente este último durante a replicação do DNA. Um exem
de análogo de bases é o 5-bromouracil (5BU), um análogo da timina, que po
Figura 1. Representação de um
fragmento de fita simples de DNA
onde está destacado um sítio
apurínico, o qual não apresenta a bas
nitrogenada no nucleotídeo.
Figura 1 – Representação de um
fragmento de fita simples de DNA onde
está destacado um sítio apurínico, o
qual não apresenta a base nitrogenada
no nucleotídeo.
152.
151
do DNA. Umexemplo de análogo de bases é o 5-bromouracil (5BU),
um análogo da timina, que pode ser incorporado em seu lugar durante
a replicação. O análogo normalmente pareia com a adenina, mas às
vezes pode parear com a guanina levando a erros nas replicações que
seguem a sua incorporação (Figura 2).
Figura 2 – Replicação do DNA com a presença do análogo de bases 5-bromouracil (5BU).
O 5BU é incorporado no lugar da timina e nas replicações seguintes ele pode parear com
guanina. Esta guanina pareará na rodada seguinte de replicação com citosina.
As mutações também podem ser induzidas pelos chamados agen-
tes alquilantes. Estas substâncias doam grupos alquila metil (CH3) e
etil (CH3-CH2) para as bases nitrogenadas, modificando seus padrões
naturais de pareamento durante a replicação do DNA. Um exemplo
de substância alquilante é o etilmetilsulfonato (EMS), o qual doa um
grupo etil para a guanina, de modo que esta forma par com timina. O
ser incorporado em seu lugar durante a replicação. O análogo normalmente
pareia com a adenina, mas às vezes pode parear com a guanina levando a
erros nas replicações que seguem a sua incorporação (Figura 2).
Figura 2. Replicação do DNA com a presença do análogo de bases 5-
bromouracil (5BU). O 5BU é incorporado no lugar da timina e nas replicações
seguintes ele pode parear com guanina. Esta guanina pareará na rodada
seguinte de replicação com citosina.
As mutações também podem ser induzidas pelos chamados agentes
alquilantes. Estas substâncias doam grupos alquila metil (CH3) e etil (CH3-
CH2) para as bases nitrogenadas, modificando seus padrões naturais de
pareamento durante a replicação do DNA. Um exemplo de substância
alquilante é o etilmetilsulfonato (EMS), o qual doa um grupo etil para a guanina,
de modo que esta forma par com timina. O EMS também adiciona um
grupamento etil na timina, provocando o seu pareamento com a guanina.
A desaminação pode ocorrer de forma espontânea, como dito
153.
152
EMS também adicionaum grupamento etil na timina, provocando o
seu pareamento com a guanina.
A desaminação pode ocorrer de forma espontânea, como dito
anteriormente, ou ser induzida. Algumas substâncias produzem desa-
minação, um exemplo é o ácido nitroso que desamina a citosina con-
vertendo-a em uracila. Durante a replicação esta uracila pareará com
a adenina. O ácido nitroso também desamina a guanina, produzindo
um nucleotídeo com a base xantina, o qual pode parear com a timina.
Outra substância que induz mutações é a Hidroxilamina, res-
ponsável por adicionar grupos hidroxila às citosinas. Esta alteração
química produz uma estrutura rara da citosina, de modo que durante
a replicação esta pareia com a adenina.
Outras substâncias que ocasionam mutações são os agentes inter-
calantes, os quais se intercalam dentro da dupla fita de DNA, causando
inserções e deleções. Exemplos de substâncias intercalantes são a
proflavina, a acridina laranja, a dioxina e o brometo de etídio (Figura 3).
Figura 3 – Dupla fita de DNA com uma molécula de Brometo de Etídio intercalada.
uracila. Durante a replicação esta uracila pareará com a adenina. O ácido
nitroso também desamina a guanina, produzindo um nucleotídeo com a base
xantina, o qual pode parear com a timina.
Outra substância que induz mutações é a Hidroxilamina, responsável
por adicionar grupos hidroxila às citosinas. Esta alteração química produz uma
estrutura rara da citosina, de modo que durante a replicação esta pareia com a
adenina.
Outras substâncias que ocasionam mutações são os agentes
intercalantes, os quais se intercalam dentro da dupla fita de DNA, causando
inserções e deleções. Exemplos de substâncias intercalantes são a proflavina,
a acridina laranja, a dioxina e o brometo de etídio (Figura 3).
Figura 3. Dupla fita de DNA com uma molécula de Brometo de Etídio
intercalada.
As mutações também podem ser geradas por radiações, exemplos são
os raios X, que podem quebrar o DNA em fita simples ou dupla e a luz
154.
153
As mutações tambémpodem ser geradas por radiações, exemplos
são os raios X, que podem quebrar o DNA em fita simples ou dupla
e a luz ultravioleta, que pode promover a formação de dímeros de
pirimidinas (pareamento de timina-timina, citosina-citosina ou timina-
citosina). Esses dímeros distorcem a dupla fita de DNA e bloqueiam a
replicação até serem consertados pelos mecanismos de reparo.
Onde Ocorrem as Mutações?
Nos organismos multicelulares podemos encontrar dois grandes
tipos de células, as somáticas e as gaméticas (germinativas). As pri-
meiras apresentam a mitose como mecanismo de divisão celular, já as
células germinativas se dividem, principalmente, por meiose. Desta
forma, dependendo do tipo celular onde ocorrem, podemos classificar
as mutações em somáticas e gaméticas. Assim, as mutações somáticas
são as que ocorrem nas células somáticas e se espalham por mitose
em um organismo. Este tipo de mutação é importante porque pode
desencadear o câncer. As mutações gaméticas surgem nas células que
originam gametas e podem ser transmitidas, podendo se expressar na
linhagem somática da próxima geração. Desta forma, as mutações em
células germinativas são muito importantes para a evolução.
Como se Classificam as Mutações?
Em relação a sua natureza molecular podemos classificar as muta-
ções em substituições, inserções e deleções. As substituições consis-
tem na troca de um nucleotídeo por outro. Podem ser de dois tipos:
Transições ou Transversões (Figura 4). Nas transições uma purina é
trocada por outra purina, ou uma pirimidina por outra pirimidina. Nas
transversões uma purina é trocada por uma pirimidina ou vice-versa.
As inserções e deleções consistem na adição ou deleção de um ou
alguns nucleotídeos. O principal problema desses tipos de mudanças
é que afetam a matriz de leitura. Os nucleotídeos do RNA mensageiro
são lidos de três em três durante a tradução. Quando adicionamos ou
deletamos nucleotídeos a matriz de leitura muda do ponto da adição
155.
154
ou deleção atéo códon de parada. Assim, este tipo de mutação poderá
mudar a sequência de aminoácidos desde o ponto onde ocorre a mu-
tação até o códon de parada da tradução. Caso a inserção ou deleção
for de três, ou múltiplos de três nucleotídeos, a mudança será menor,
podendo afetar somente os códons que perderam ou ganharam nu-
cleotídeos, mas não se estenderá até o códon de parada da tradução.
Figura 4 – No diagrama são mostradas as transições (A para G e C para T) e as
transversões (A para C e vice-versa, A para G e vice-versa, C para A e vice-versa, e C
para G e vice-versa) das bases púricas (A e G) e pirimidínicas (T e C).
As mutações ocorrem ao nível molecular e nem sempre apresen-
tam efeitos fenotípicos. Quando não ocorre mudança no fenótipo,
a mutação causa apenas uma alteração no nucleotídeo e não gera
uma mudança no aminoácido da proteína. Isso se deve ao fato de que
vários aminoácidos são codificados por mais de um códon (código
genético degenerado). As mutações que mudam os nucleotídeos e
não mudam o aminoácido são chamadas de silenciosas (Tabela 1). A
Figura 4. No diagrama são mostradas as transições (A para G e C
transversões (A para C e vice-versa, A para G e vice-versa, C pa
versa, e C para G e vice-versa) das bases púricas (A e G) e pirim
C).
As mutações ocorrem ao nível molecular e nem sempre
efeitos fenotípicos. Quando não ocorre mudança no fenótipo, a mu
apenas uma alteração no nucleotídeo e não gera uma mudança no
da proteína. Isso se deve ao fato de que vários aminoácidos são
por mais de um códon (código genético degenerado). As mutações
156.
155
seguir classificaremos asmutações de acordo com suas consequências
sobre o fenótipo.
Tabela 1 – Sequência de DNA para o tipo selvagem e para as mutações gênicas
silenciosas, não silenciosas e de parada, bem como seus respectivos produtos de
RNA mensageiro e o aminoácido codificado.
Tipo
selvagem
Mutação gênica
DNA
Silenciosa Não Silenciosa De parada
TTC TTT TCC ATC
RNA mensageiro AAG AAA AGG UAG
Aminoácido Lisina Lisina Arginina
Códon de
Parada
O efeito fenotípico da mutação é observado na comparação de
um indivíduo mutante com outro individuo sem a mutação, chamado
de selvagem (Figura 5). Quando observamos que o fenótipo mutante
é diferente do selvagem a mutação é chamada de direta. Ás vezes
o mutante muta novamente e restaura o fenótipo selvagem. Neste
caso a nova mutação recupera o nucleotídeo original e restaura o
mesmo aminoácido. Há também as mutações supressoras, neste caso
a mudança não reverte o estado molecular selvagem, mas esconde ou
suprime o efeito fenotípico da mutação direta. A mutação supressora
não anula o que aconteceu na primeira mutação, mas, devido a uma
segunda mutação, os efeitos da primeira são anulados.
Figura 5. Drosophila melanogaster. A esquerda uma mosca com fenótipo
mutante para a cor de olhos e a direita o fenótipo selvagem.
MECANISMOS DE REPARO
Figura 5 – Drosophila melanogaster. A
esquerda uma mosca com fenótipo mutante
para a cor de olhos e a direita o fenótipo
selvagem.
157.
156
As mutações desubstituição também podem ter um efeito
fenotípico se causarem mudanças no aminoácido, sendo chamadas
de mutações não silenciosas (Tabela 1). Ás vezes o novo aminoá-
cido não afeta a função da proteína, nesse caso a substituição
não silenciosa é considerada neutra. Outras vezes a mutação
não silenciosa leva a perda da função da proteína, nesse caso a
mutação é chamada de perda de função. Há também mutações
não silenciosas onde é gerada uma nova função para um gene,
são as mutações de ganho de função.
A mudança de nucleotídeo também pode originar um códon
de parada antes do observado na forma selvagem, nesse caso
essa substituição é chamada de mutação de parada (Figura 5).
Outras mutações com efeito fenotípico são as condicionais,
nas quais o fenótipo mutante aparece em determinadas con-
dições, por exemplo, a temperaturas acima de 28°C ocorre o
fenótipo mutante, mas abaixo dessa temperatura se expressa o
fenótipo selvagem.
Finalmente, temos as mutações letais, onde o indivíduo não
nasce devido à presença de uma mutação.
Mecanismos de Reparo
Nosso DNA está constantemente sofrendo mudanças, tanto de
forma espontânea como por agentes físicos, como as radiações,
ou substâncias químicas. Mesmo assim, esta molécula é muito
estável, já que conta com sistemas que reparam a grande maioria
das mutações. Existe um grande número de sistemas de reparo,
mas a maioria deles requer que o DNA seja de fita dupla. Assim,
uma das fitas será utilizada como molde para fazer o reparo. Outra
característica importante é que o mesmo erro pode ser consertado
por várias vias de mecanismos de reparo, mostrando assim sua
eficiência. Poucas são as mutações que escapam dos sistemas de
reparo. Existem vários mecanismos que reparam danos no DNA,
a seguir descreveremos alguns dos principais.
158.
157
Reparo de PareamentoErrado
A replicação do DNA é um mecanismo quase perfeito, porém em
baixa frequência acontecem erros durante este processo. Quando
existempareamentosdebasesdiferentesdeAdenina-TiminaeGuanina-
Citosina ocorre uma deformação da dupla fita de DNA. Nesta situação,
um conjunto de enzimas detecta essas mudanças estruturais na mo-
lécula de DNA. Após o reconhecimento do erro, as enzimas eliminam
o trecho distorcido e preenchem o espaço com novos nucleotídeos,
usando o filamento de DNA como molde. Porém, qual das duas fitas
de DNA deve ser utilizada como molde, ou, em outras palavras, como
saber qual a fita de DNA contém a base mal incorporada? Para tal, esse
sistema de reparo verifica qual das fitas apresenta bases metiladas. Na
fita mais recentemente sintetizada, a que contém o erro (mutação),
a metilação ainda não ocorreu. Portanto, o reconhecimento da fita
metilada garante o reparo correto da base erroneamente incorporada.
Reparo Direto
Neste sistema de reparo os nucleotídeos alterados são corrigidos
para sua estrutura original. Por exemplo, a luz UV gera dímeros de
pirimidinas que impedem a continuidade da replicação. Em E. coli
existe uma enzima, chamada de Fotoliase, que absorve a energia da
luz e quebra as ligações covalentes presentes nos dímeros, permitin-
do o prosseguimento da replicação. Este sistema de reparo se chama
fotorreativação enzimática (Figura 6).
Reparo por Excisão de Bases
É comum que algumas bases sejam modificadas com o tempo. O
reparo por excisão de nucleotídeos é responsável por restaurar essas
bases nitrogenadas ao seu estado inicial. As bases modificadas (mu-
tadas) são eliminadas por um grupo de enzimas chamadas de DNA
glicosilases. Neste sistema cada enzima reconhece e elimina um tipo
de base modificada, mediante a ruptura da ligação covalente entre a
base e o açúcar. Depois da remoção das bases uma enzima chamada
159.
158
de AP (apurínicaou apirimidínica) corta a ligação fosfodiéster e outra
enzima remove o açúcar (desoxirribose). A DNA polimerase adiciona
novos e corretos nucleotídeos no extremo 3’ OH exposto e a DNA
ligase catalisa as novas ligações fosfodiéster (Figura 7).
DNA polimerase adiciona novos e corretos nucleotídeos no extremo 3’ OH
exposto e a DNA ligase catalisa as novas ligações fosfodiéster (Figura 7).
Figura 6. Esquema do mecanismo de reparo por fotorreativação enzimática.
Figura 6 – Esquema do mecanismo de reparo por fotorreativação enzimática.
160.
159
Figura 7 –Esquema do mecanismo de reparo por excisão de bases.
Figura 7. Esquema do mecanismo de reparo por excisão de bases.
161.
160
Reparo por Excisãode Nucleotídeos
Este tipo de reparo atua sobre diferentes danos no DNA geral-
mente de extensões maiores, que causam uma distorção na dupla fita
desta molécula. O primeiro passo do sistema de reparo por excisão
de nucleotídeo é feito por um conjunto de enzimas que revisam o
DNA a procura de distorções na dupla fita. Quando um problema é
encontrado um outro grupo de enzimas separa o DNA em fitas simples
na região mutada e um conjunto de enzimas estabiliza o DNA como
fita simples. A fita que apresenta a mutação é removida e o espaço é
preenchido pela DNA polimerase e ligado pela DNA ligase (Figura 8).
Figura 8 – Esquema do mecanismo de reparo por excisão de nucleotídeos.
155
REPARO POR EXCISÃO DE NUCLEOTÍDEOS
Este tipo de reparo atua sobre diferentes danos no DNA geralmente de
extensões maiores, que causam uma distorção na dupla fita desta molécula. O
primeiro passo do sistema de reparo por excisão de nucleotídeo é feito por um
conjunto de enzimas que revisam o DNA a procura de distorções na dupla fita.
Quando um problema é encontrado um outro grupo de enzimas separa o DNA
em fitas simples na região mutada e um conjunto de enzimas estabiliza o DNA
como fita simples. A fita que apresenta a mutação é removida e o espaço é
preenchido pela DNA polimerase e ligado pela DNA ligase (Figura 8).
Figura 8. Esquema do mecanismo de reparo por excisão de nucleotídeos.
162.
161
EXERCÍCIOS
1 – Todasas mutações são adaptativas? Justifique sua resposta.
2 – Diferencie mutações espontâneas de mutações induzidas.
3 – “Cometer erros é a chave para o progresso. Há momentos em que é
importante não cometer erro algum – pergunte a qualquer cirurgião
ou piloto de avião. No entanto (…) os erros não são apenas oportuni-
dades valiosas para aprendermos; eles são, de forma significativa, a
única oportunidade para aprendermos algo relativamente novo. (…) A
evolução biológica se dá através de uma grande e inexorável sequência
de tentativas e erros – e sem os erros as tentativas não teriam levado a
nada.”(Adaptado de DENNETT, Daniel C. In: BROCKMAN, J. e MATSON
K. As coisas são assim. São Paulo: Cia. das Letras, 1997)
O processo biológico que se relaciona com o texto apresentado pelo autor é:
a) o reparo das lesões gênicas.
b) a indução de mutações programadas.
c) a geração de organismos transgênicos.
d) a alteração aleatória de nucleotídeos do DNA.
e) a clonagem genética.
4 – As mutações podem apresentar importância evolutiva quando ocorrem
em células:
a) diploides.
b) somáticas.
c) germinativas.
d) embrionárias.
e) anucleadas.
163.
162
5 – Podeser entendida como alterações do código bases nitrogenadas do
DNA, que originam novas versões de genes:
a) a evolução.
b) a mutação.
c) o estado homozigoto.
d) a mitose.
e) a fecundação.
6 – As mutações podem ser induzidas por agentes físicos e químicos, deno-
minados genericamente de agentes:
a) gênicos.
b) mutagênicos.
c) alélicos.
d) evolutivos.
e) radioativos.
7 – Quando uma mutação leva à substituição de um códon por outro que
significa o mesmo aminoácido, a mutação resultante é conhecida como:
a) adição.
b) transição.
c) transversão.
d) deleção.
e) silenciosa.
8 – A substituição de uma base nitrogenada por outra na molécula de DNA:
a) sempre resultará em uma nova proteína que difere da antiga pelo
número de aminoácidos.
b) sempre resultará em uma nova proteína que difere da antiga pela
ordenação dos aminoácidos.
c) sempre resultará em uma nova proteína que difere da antiga pela
troca de três aminoácidos.
164.
163
d) sempre resultaráem nova proteína que difere da antiga pelo menor
tamanho.
e) pode resultar em uma nova proteína, com a mesma ordem de aminoá-
cidos ou não, e até mesmo com um tamanho distinto da proteína original.
9–Atualmentesãobemconhecidososefeitosadversosàsaúdehumanacausa-
dospordiversospoluentesambientais,especialmenteaquelesquepossuem
potencialidades mutagênicas ou carcinogênicas, os quais, devido à sua
interação com mecanismos genéticos, podem causar mutações e doenças
nas gerações futuras. Sobre este assunto observe as seguintes afirmações:
I. Mutações são modificações do material genético que podem ser trans-
mitidas à prole (descendência ou células filhas).
II. A mutação pode ser espontânea ou induzida por agentes físicos,
químicos ou biológicos com potencial mutagênico.
III. Mutação é toda alteração do material genético que resulta sempre
de segregação ou recombinação cromossômicas.
IV. Mutações gênicas podem ser causadas por poluentes ambientais e
provocar alterações responsáveis pelo aparecimento de genótipos
diferentes numa população.
A alternativa que contém as afirmações corretas é:
a) I e III.
b) II e III.
c) I, II e IV.
d) IV e III.
e) III.
10 – O câncer é uma doença causada pelo aumento descontrolado do número
de células em uma ou algumas partes de um organismo, devido a muta-
ções no material genético. Por essa razão podemos afirmar que o câncer:
a) é uma doença contagiosa.
b) é uma doença genética.
165.
164
c) é umadoença incurável.
d) afeta os genes envolvidos na síntese de ATP.
e) somente será curado com o sequenciamento do genoma humano.
11 – Entre os efeitos causados por radiações está a ocorrência de mutações
no DNA. Sobre o processo de mutação é incorreto afirmar que:
a) é sempre irreversível.
b) pode ser causada por agentes biológicos.
c) pode ser causada por agentes químicos.
d) pode ser causada por agentes físicos.
e) pode ser “espontânea”
12 – Observe as cinco afirmações abaixo sobre o assunto mutações gênicas:
I. As mutações podem ocorrer tanto em células somáticas como em
células germinativas.
II. Somente as mutações ocorridas em células somáticas poderão produzir
alterações transmitidas às próximas gerações, independentemente
do sistema reprodutivo.
III. Apenas as mutações que atingem as células germinativas da espécie
humana podem ser transmitidas aos descendentes.
IV. As mutações não podem ser “espontâneas”, mas apenas causadas por
fatores mutagênicos, tais como agentes químicos e físicos.
V. As mutações são fatores importantes na promoção da variabilidade
genética e para a evolução das espécies.
Assinale a alternativa que contém todas as afirmações corretas.
a) I, II e III.
b) I, III e V.
c) I, IV e V.
d) II, III e IV.
e) II, III e V.
166.
165
REFERÊNCIAS BIBLIOGRÁFICAS
ALBERTS, B.;BRAY, D.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K. &
WALTER, P. Fundamentos de Biologia Celular. Ed. Artes Médicas, São Paulo.
1999.
BROWN, T.A. Genética, um enfoque molecular. 3ª Ed. Rio de Janeiro: Editora
Guanabara Koogan S.A., 1999
DE ROBERTIS, E.; HIB, J. Bases da Biologia Celular e Molecular. Ed. Guanabara
Koogan S.A. 4ª ed. Rio de Janeiro/RJ, 389p. 2006.
GRIFFITHS, A.J.F. et al. Introdução à Genética. 7ª edição. Rio de Janeiro: Ed.
Guanabara Koogan, 2002
JUNQUEIRA, L. C. & CARNEIRO, J. Biologia Celular e Molecular. Ed. Guanabara
Koogan S.A. 8ª ed. Rio de Janeiro/RJ, 2005.
PIERCE, B.A. Genética: Um enfoque conceitual. Rio de Janeiro: Guanabara
Koogan, 2004.
SNUSTAD, D.P. e SIMMONS, M.J. Fundamentos de genética. 2° ed. Rio de
Janeiro: Ed. Guanabara Kogan, 2001
167.
166
INFORMAÇÕES DOS AUTORES
Coordenadorado Projeto:
Maria Mascena Diniz Maia
Professora Titular da Universidade Federal Rural de Pernambuco
Doutora em Ciências Biológicas pela Universidade Federal de Pernambuco
Mestre em Bioquímica e Fisiologia pela Universidade Federal de Pernambuco
Bacharel em Farmácia pela Universidade Federal de Pernambuco
Demais autores:
Paulo Roberto Eleutério de Souza
Professor Associado da Universidade Federal Rural de Pernambuco
Doutor em Ciências Biológicas pela Universidade Federal de Pernambuco
Mestre em Genética pela Universidade Federal de Pernambuco
Bacharel em Farmácia pela Universidade Federal de Pernambuco
Hildson Dornelas Angelo da Silva
Professor Adjunto do Instituto Federal de Pernambuco
Doutor em Genética pela Universidade Federal de Pernambuco
Mestre em Biologia Celular e Molecular Aplicada pela Universidade de Pernambuco
Bacharel em Ciências Biológicas pela Universidade de Pernambuco
Licenciado em Ciências Biológicas pela Universidade Federal Rural de Pernambuco
Fernanda Cristina Bezerra Leite
Professora Adjunta da Universidade Federal Rural de Pernambuco
Doutora em Ciências Biológicas pela Universidade Federal de Pernambuco
Mestre em Genética pela Universidade Federal de Pernambuco
Bacharel em Ciências Biológicas pela Universidade Federal de Pernambuco
168.
167
Ana Cristina LauerGarcia
Professora Associada da Universidade Federal de Pernambuco
Doutora em Genética e Biologia Molecular pela Universidade Federal do Rio
Grande do Sul
Mestre em Genética e Biologia Molecular pela Universidade Federal do Rio
Grande do Sul
Licenciada em Ciências Biológicas pela Universidade Federal do Rio Grande do Sul
Martín Alejandro Montes
Professor Adjunto da Universidade Federal Rural de Pernambuco
Doutor em Genética e Biologia Molecular pela Universidade Federal do Rio Grande
do Sul
Mestre em Genética e Biologia Molecular pela Universidade Federal do Rio Grande
do Sul
Licenciado em Genética pela Universidad Nacional de Misiones



![Profª. Maria José de Sena — Reitora
Prof. Marcelo Brito Carneiro Leão — Vice-Reitor
Conselho Editorial
Presidente: Marcelo Brito Carneiro Leão; Diretor da Editora da UFRPE: Bruno de Souza
Leão; Diretora do Sistema de Bibliotecas da UFRPE : Maria Wellita Santos
Conselheiros: Álvaro José de Almeida Bicudo, Fernando Joaquim Ferreira Maia
Maria do Rosário de Fátima Andrade, Monica Lopes Folena Araújo, Rafael Miranda
Tassitano, Renata Pimentel Teixeira
Revisão final dos autores
Editora filiada à
Positivo e negativo
Manual de Identidade Visual ABEU 07
Todos os direitos reservados. É proibida a reprodução total ou parcial do trabalho sem autorização da
universidade federal rural de pernambuco - departamento de biologia - genética e dos autores.
Ficha catalográfica
G328 Genética geral para universitários / Maria de Mascena Diniz
Maia, coordenadora; Paulo Roberto Eleutério de Souza ...
[et al]. -- 1. ed. -- Recife : EDUFRPE, 2015.
147 p. : il.
ISBN : 978-85-7946-229-0
Referências.
1. Genética I. Maia, Maria de Mascena Diniz, coord.
II. Souza, Paulo Roberto Eleutério de
CDD 575.1](https://image.slidesharecdn.com/livrogeneticageralweb-250318180658-5248729d/85/livro_geneticageralweb-pdfccccccccccccccc-3-320.jpg)