

Recomendados
Mais conteúdo relacionado
Destaque
Destaque (20)
Semelhante a Présent i cours1
Présent i cours1
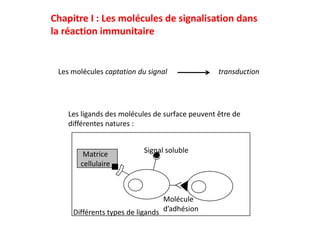
- 1. Chapitre I : Les molécules de signalisation dans la réaction immunitaire Les molécules captation du signal transduction Les ligands des molécules de surface peuvent être de différentes natures : Matrice Signal soluble cellulaire Molécule Différents types de ligands d’adhésion
- 2. •Les molécules de la surface cellulaire •Aspects cellulaires. La superfamille des Ig Les récepteurs des Ig (RFc : CD16, CD32, CD64), les molécules associées aux TCR (CD3, CD4 et CD8) et aux BCR (CD19, CD79). Les molécules d’activation et d’inhibition des NK qui interagissent avec les molécules HLA de classe I. Les intégrines (exemple la LFA1, leukocyte function antigen, CD11a/CD18) sont des glycoprotéines membranaires dont la fonction est l’adhésion à d’autres cellules et à la matrice extracellulaire. Les sélectines sont des lectine essentielles pour l’adhésion par leurs actions de ralentissement du flux cellulaire essentiellement au niveau vasculaire. Elles sont de 3 types selon leurs expressions : endothéliale CD62E, leucocytaire CD62L et plaquettaire CD62P. Leurs ligants sont oligosaccharides GlyCAM-1 et CD34. Les récepteurs des cytokines sont des molécules différentes selon la cytokine captée. Per exemple le R TNF est le récepteur de TNF. Les récepteurs des cytokines sont membranaires, ils peuvent être solubles.
- 6. •Aspects moléculaires Les gènes codant pour les molécules de surface présentent : un exon codant pour le peptide signal (L) et d’autres codant pour la partie cytoplasmique, (IC) transmembranaire (TM) et extracellulaire (EC). L EC EC TM IC Poly A L EC EC TM IC L EC Poly A
- 7. •Fonctions des molécules de surface •La captation du ligand Elle est réalisée grâce aux domaines extracellulaires de la molécule de surface. La spécificité de ces molécules pour leurs ligands peut être déterminée par des séquences peptidiques et par leurs structures tridimensionnelles. •Induction du signal Elle est réalisée par le domaine intracellulaire des molécules de surface. Ces domaines peuvent avoir une activité enzymatique tells que les motifs ITAM (D/ExxYxxL) (immunoreceptor tyrosine based activation motif). Ces motifs sont trouvés au niveau des immuns récepteurs (CD3 des LT, CD79 des LB) et sur le récepteur R Fc des Ig, CD5 ou le DAP12 des NK. La mise en contact des motifs ITAM et du site SH2 de l’enzyme thyrosine kinase induit la phosphorisation des tyrosines des motifs ITAM est le point de départ de l’activation de la cellule. Après phosphorilation du motif ITAM, une molécule ZAP-70 est activée et active à son tour d’autres substrats cellulaire. Une activité antagoniste (une inhibition) est assurée par des motifs dit ITIM (immunoreceptor tyrosine-based inhibition motives) interagissent avec une tyrosine phosphatase (PTP ou CD45) qui abouti à déphosphorisation des tyrosines de diverses molécules et induit l’inhibition du signal.