molecular biology presentation.pptx
•Transferir como PPTX, PDF•
1 gostou•16 visualizações

DNA , Structure, replication

Recomendados
Mais conteúdo relacionado
Semelhante a molecular biology presentation.pptx
Semelhante a molecular biology presentation.pptx (20)
Último
Último (20)
molecular biology presentation.pptx
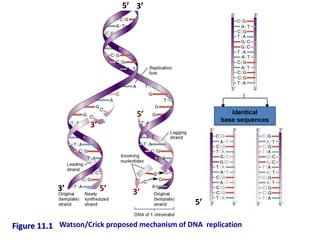
- 1. Figure 11.1 Identical base sequences 5’ 5’ 3’ 3’ 5’ 5’ 3’ 3’ Watson/Crick proposed mechanism of DNA replication
- 2. • In the late 1950s, three different mechanisms were proposed for the replication of DNA – Conservative model • Both parental strands stay together after DNA replication – Semiconservative model • The double-stranded DNA contains one parental and one daughter strand following replication – Dispersive model • Parental and daughter DNA are interspersed in both strands following replication Proposed Models of DNA Replication
- 3. Three models for DNA replication
- 4. • Matthew Meselson and Franklin Stahl experiment in 1958 – Grow E. coli in the presence of 15N (a heavy isotope of Nitrogen) for many generations • Cells get heavy-labeled DNA – Switch to medium containing only 14N (a light isotope of Nitrogen) – Collect sample of cells after various times – Analyze the density of the DNA by centrifugation using a CsCl gradient
- 5. CsCl Density Gradient Centrifugation 15N 14N DNA
- 7. Interpreting the Data After one generation, DNA is “half-heavy” After ~ two generations, DNA is of two types: “light” and “half-heavy” This is consistent with only the semi-conservative model
- 9. Three main features of the DNA synthesis reaction: 1. DNA polymerase I catalyzes formation of phosphodiester bond between 3’-OH of the deoxyribose (on the last nucleotide) and the 5’-phosphate of the dNTP. • Energy for this reaction is derived from the release of two of the three phosphates of the dNTP. 2. DNA polymerase “finds” the correct complementary dNTP at each step in the lengthening process. • rate ≤ 800 dNTPs/second • low error rate 3. Direction of synthesis is 5’ to 3’
- 10. DNA polymerase Comparison of the properties of the DNA polymerase of E.Coli Pol I Pol II Pol III 5’ → 3’ Polymerase Yes Yes Yes 3’ → 5’ Exonuclease Yes Yes Yes 5’ → 3’ Exonuclease Yes No No Structure Polypeptide Poly peptide Multimeric complex Function Repair, Primer excision Error Prone repair polymerase (SOS inducible) Principle replication polymerase Subunits of E.Coli Pol III holoenzyme Core Subunit α 5’→ 3’ Polymerase activity, required for DNA synthesis ε 3’→ 5’ exonuclease activity, required for proofreading Q Function unknown. Accessory τ DNa dependent ATPase, required for initiation. γ DNA dependent ATPase forming γ complex (with 4 peptides) facilitates β subunit binding. δ, δ1, χ, Ψ Forms γ complex required for loading & unloading β subunit β ‘Sliding clamp’, forms preinitiation complex with DNA a process which requires ATP dependent activity of the γ complex.
- 11. EUKARYOTIC DNA POLYMERASES Mammalian Name α β γ δ ε Yeast Name Pol 1 Pol 4 Pol M Pol 3 Pol 2 Yeast gene POL 1 POL 4 MIP 1 POL3 POL2 Location Nuclear Nuclear Mitochondria l Nuclear Nuclear No. of subunit 4 1 2 2 >1 5’→3’Polymerase Yes Yes Yes Yes Yes 3’→ 5’ Exonuclease No No Yes Yes Yes Primase Yes No No No No Associated facotors None None None PCNA None Processivity Moderat e Low High High with PCNA High Function Lagging strand priming Repair polymerase Organelle polymerase Principle replicative polymerase Unknown
- 12. Issues to Resolve for DNA Replication to Occur • Helix must be unwound • Duplex over winding issues due to fork migration • Primer synthesis • One fork, two antiparallel strands… • Primer removal • Connecting the pieces… • Proofreading and error correction
- 13. • Initiation of replication, major elements: Segments of single-stranded DNA are called template strands. Gyrase (a type of topoisomerase) relaxes the supercoiled DNA. Initiator proteins and DNA helicase binds to the DNA at the replication fork and untwist the DNA using energy derived from ATP (adenosine triphosphate). (Hydrolysis of ATP causes a shape change in DNA helicase) DNA primase next binds to helicase producing a complex called a primosome (primase is required for synthesis), Primase synthesizes a short RNA primer of 10-12 nucleotides, to which DNA polymerase III adds nucleotides. Polymerase III adds nucleotides 5’ to 3’ on both strands beginning at the RNA primer. The RNA primer is removed and replaced with DNA by polymerase I, and the gap is sealed with DNA ligase. Single-stranded DNA-binding (SSB) proteins (>200) stabilize the single-stranded template DNA during the process.
- 14. • DNA synthesis begins at a site termed the origin of replication • Each bacterial chromosome has only one • Synthesis of DNA proceeds bidirectionally around the bacterial chromosome – eventually meeting at the opposite side of the bacterial chromosome • Where replication ends BACTERIAL REPLICATION
- 15. Unwinding the Helix • E. coli origin called oriC – 245 bp region with repeating 9mer and 13mer repeating sequences • Each 9mer bound by several DnaA protein monomers • DnaB and DnaC then bind – Form a helicase that uses ATP to open helix • Ssb protein binds to ss DNA • Topoisomerase DNA gyrase eliminates positive supercoiling caused by unwinding (uses ATP)
- 19. Initiation of Synthesis • DNAPs cannot start new polynucleotide strand – Must add on to something • Primase (specialized RNAP) makes a short (5-15 nucleotide) RNA primer – 3’end is used by DNAP III for new strand initiation • RNA primer must later be removed and replaced by DNA – Done by DNAP I using 5’ exonuclease activity
- 20. RNA Primers for Initiation • RNA primers for DNA synthesis are fairly universal in their use – E. coli, mammals, viruses/phage – But not all viruses…
- 21. Initiation of Replication at oriC • DNA replication is initiated by the binding of DnaA proteins to the DnaA box sequences – causes the region to wrap around the DnaA proteins and separates the AT-rich region
- 22. Uses energy from ATP to unwind the duplex DNA SSB SSB SSB SSB
- 23. • DNA helicase separates the two DNA strands by breaking the hydrogen bonds between them • This generates positive supercoiling ahead of each replication fork – DNA gyrase travels ahead of the helicase and alleviates these supercoils • Single-strand binding proteins bind to the separated DNA strands to keep them apart • Then short (10 to 12 nucleotides) RNA primers are synthesized by DNA primase – These short RNA strands start, or prime, DNA synthesis
- 24. Fig. 11.9a(TE Art) Able to covalently link together Unable to covalently link the 2 individual nucleotides together 5’ 5’ 5’ 5’ 5’ 5’ 3’ 3’ 3’ 3’ 3’ DNA Polymerase Cannot Initiate new Strands
- 26. DNA Polymerase III- does the bulk of copying DNA in Replication
- 27. Figure 11.8 Schematic representation of DNA Polymerase III Structure resembles a human right hand Template DNA thread through the palm; Thumb and fingers wrapped around the DNA
- 28. Two dimensional view of a replication fork Direction of synthesis on leading strand 3’ 5’ 3’ 5’ 3’ 5’
- 29. Figure 11.13 “Three Dimensional” view of Replication Fork Direction of fork movement Direction of synthesis Of lagging strand Direction of synthesis of leading strand
- 30. Proofreading by the 3’ 5’ exonuclease activity of DNA polymerases during DNA replication.
- 31. DNA Duplexes Have Antiparallel Strands… • Replication forks move off from origins – But to replicate both strands one DNAP or replisome would seem to need to move back toward the origin… • Efforts to find a 3’ to 5’ DNAP for the “other” strand failed • One strand is made in a discontinuous fashion (discontinuous or lagging strand)
- 32. Leading and Lagging Strands • Discontinuous synthesis discovered by Okazaki • Pulse-chase experiment revealed that new DNA was initially small pieces but became very large with time • Right conclusion from wrong results…
- 33. Lagging Strand Synthesis • Lagging strand made as a series of 1000-2000 nucleotide pieces (bacteriophage), each begging with an RNA primer (Okazaki fragments) • Primers removed by nick translation action of DNAP I • Fragments connected by DNA ligase – Using NAD+ (E. coli) or ATP (eukaryotes and most others)
- 34. Concurrent Synthesis of Leading and Lagging Strands • Kornberg model – Process should be processive, not distributive – Makes no sense for DNAP molecules to move away from the fork and then have to return – Has two DNAP III core enzymes connected to each other • One synthesizes each strand – One continuously and one using a looping discontinuous method that produces short Okazaki fragments
- 35. 3 Polymerase III Leading strand base pairs 5’ 5’ 3’ 3’ Supercoiled DNA relaxed by gyrase & unwound by helicase + proteins: Helicase + Initiator Proteins ATP SSB Proteins RNA Primer primase 2 Polymerase III Lagging strand Okazaki Fragments 1 RNA primer replaced by polymerase I & gap is sealed by ligase
- 36. Fig. 3.8 Model of DNA Replication
- 37. Connecting Leading and Lagging Strand Synthesis Polymerase dimer synthesizes both strands simultaneously using both a continuous and a looping discontinuous approach
- 38. Activities at the Replication Fork • DNAP I and DNAP III, helicase (DnaBC, Rep), DNA gyrase, SsB, primase, DNA ligase
- 39. Figure 11.4 Overview of bacterial DNA replication
- 40. Autoradiography: Radioactivity darkens film Radioactive bacterial colonies on an agar petri dish
- 41. Directionality of the DNA strands at a replication fork Leading strand Lagging strand Fork movement
- 42. Nicks are single strand breaks in double stranded DNA
- 43. Proofreading • DNAPs have 3’exonuclease activity specific for single-stranded DNA – Unpaired nucleotides • DNAPs “back up” to remove unpaired nucleotides and then add correct base – Improves fidelity of replication by 100X – In E. coli activity in the e subunit of DNAP III
- 44. • DNA polymerases can only synthesize DNA only in the 5’ to 3’ direction and cannot initiate DNA synthesis • These two features pose a problem at the 3’ end of linear chromosomes Figure 11.24 Problem at ends of eukaryotic linear Chromosomes
- 45. • If this problem is not solved – The linear chromosome becomes progressively shorter with each round of DNA replication • The cell solves this problem by adding DNA sequences to the ends of chromosome: telomeres – Small repeated sequences (100-1000’s) • Catalyzed by the enzyme telomerase • Telomerase contains protein and RNA – The RNA functions as the template – complementary to the DNA sequence found in the telomeric repeat • This allows the telomerase to bind to the 3’ overhang
- 46. Telomerase • RNA-dependent DNA polymerase – Has its own RNA template complementary to the telomeric repeat sequence – Adds on additional ss repeat units • Unusual hairpin structure forms new primer to make ds • Cleavage of hairpin yields lengthened telomere
- 47. 11-80 Figure 11.25 Step 1 = Binding Step 3 = Translocation The binding- polymerization- translocation cycle can occurs many times This greatly lengthens one of the strands The complementary strand is made by primase, DNA polymerase and ligase RNA primer Step 2 = Polymerization
- 48. DNA Repair mechanisms • This section will review: –The Role of DNA damage and its repair in the generation of genetic diversity in bacteria –The mechanistic links of repair with recombination systems covered earlier • You should be able to discuss the effect of environment on damage and repair. For example: –Chemical and Radiation effects –Phagocytic damage in relation to pathogen survival in the host –Errors in replication and their repair.
- 49. How can DNA become damaged? • Mismatched bases – Polymerase error rate about 1 in 104 (see later lectures) – Deamination of C to U leading to mismatch • Missing bases. Hydrolysis of purine-deoxyribose bond leading to AP-site. • Structural damage. Dimer formation. • Broken phosphodiester bonds. Chemicals/radiation • REPAIR MECHANISMS NECESSARY FOR SURVIVAL.
- 50. Types of DNA Damage Summarised G A C T ds DNA Break Mismatch Thymidine dimer AP site Covalent X-linking ss Break C-U deamination
- 51. General repair mechanisms needed • EITHER Reverse damage (e.g. PHOTOREACTIVATION) • OR excise DNA and patch repair the region Photoreactivation: Discovered in Actinomycetes in 1949 UV - DNA Damage - Cell Death UV - Bright visible light - survival ! 3 Steps: •Photolyase (encoded by phrA and phrB genes in E. coli) recognises distortion at dimer. •Light activates photolyase •Dimer cleaved
- 52. General repair mechanisms needed: Excision repair. • Discovered first as a general mechanism in 1964 • T- Phage HOST CELL REACTIVATION T-Phage suspension DNA Damage Repair - WT Repair Plaques ! UV
- 54. Other repair routes. •Excision repair involves up to 20 nucleotides •uvrA,BC (D) mutants very senstive to UV light •Mismatch repair •A from of excision repair. Dam methylase involved •see later re: methylation •N-glycolylase excision repair •Uracil either misincorporated OR C deaminated to U •Uracil N-glycosylase action TO give AP site •AP endonuclease cut •Patch repairs above
- 55. How does UV light cause mutations? Discovery of error-prone repair. • RecA required for high level of UV mutagenesis •UV dose of 2 µJ/mm2 leads to 120 dimers •Long patch Error-prone repair •Post dimer initiation •Trans dimer synthesis The SOS Hypothesis Radman 1974 originally proposed an inducible repair system Requires RecA and a regulator system
- 56. SOS System in E.coli • Repair normally at low level • lexA gene identified as a regulator • Recombine normally • But NO increased UV mutagenesis (ie 30 dimers produces no extra mutants). Higher doses required • LOW DOSE - Error-free repair • HIGH DOSE - Error repair INDUCED • LexA is an autoregulated repressor • Represses level of activity of many genes • Collectively called DNA Inducible (din) genes • Includes uvrA,B,C,D and sfi etc... • RecA protease activity; Cleaves LexA • Also CI repressor inducing lysis
- 57. SOS System in E.coli PO lexA PO recA PO din PO din PO din PO din PO din PO din PO din PO din PO din PO din Low level expression PO lexA PO recA HIGH level expression
- 58. Post Replication / Recombination Repair •recA mutants VERY UV sensitive •uvrA similarly •uvrA recA mutants VERY VERY UV sensitive •recA mutants recover slowly in the dark •uvrA mutants do not UV dose Dimers/ Genotype Phenotype µJ/mm2 genome WT WT 5.0 3200 uvrA No excision repair 0.8 50 recA No recombination 0.3 20 recA, uvrA Neither 0.02 1
- 59. Post Replication / Recombination Repair Stephen C. Kowalczykowski (2000) Initiation of genetic recombination and recombination-dependent replication. TIBS 25 – April 2000 • Double-stranded-break repair – see transposition of Tn10 later • Recombination-dependent replication • Replication-dependent recombination SEE ALSO Key Reference in NATURE; STRUCTURE OF RecBCD complex • Nature 11th Nov 2004 vol 432, 187-193
- 60. RNA polymerase in bacteria In bacteria, the same enzyme catalyzes the synthesis of three types of RNA: mRNA, rRNA and tRNA. RNAP is a relatively large molecule. The core enzyme has 5 subunits (~400 kDa): • α2: the two α subunits assemble the enzyme and recognize regulatory factors. • β: this has the polymerase activity (catalyzes the synthesis of RNA). • β’: binds to DNA (nonspecifically). • ω: function not known clearly. RNA polymerase in eukaryotes Eukaryotes have several types of RNAP: • RNA polymerase I synthesizes a pre-rRNA 45S, which matures into 28 S, 18S and 5.8S rRNAs which will form the major RNA sections of the ribosome. • RNA polymerase II synthesizes precursors of mRNAs and most snRNA. This is the most studied type, and due to the high level of control required over transcription a range of transcription factors are required for its binding to promoters. For detail of RNA polymerase function please see RNA polymerase II. • RNA polymerase III synthesizes tRNAs, rRNA 5S and other small RNAs found in the nucleus and cytosol. Other RNA polymerase types in mitochondria and chloroplasts.
- 61. THE lac OPERON © 2007 Paul Billiet ODWS
- 62. The control of gene expression • Each cell in the human contains all the genetic material for the growth and development of a human • Some of these genes will be need to be expressed all the time • These are the genes that are involved in of vital biochemical processes such as respiration • Other genes are not expressed all the time • They are switched on an off at need © 2007 Paul Billiet ODWS
- 63. Operons • An operon is a group of genes that are transcribed at the same time. • They usually control an important biochemical process. • They are only found in prokaryotes. © NobelPrize.org Jacob, Monod & Lwoff © 2007 Paul Billiet ODWS
- 64. The lac Operon The lac operon consists of three genes each involved in processing the sugar lactose One of them is the gene for the enzyme β- galactosidase This enzyme hydrolyses lactose into glucose and galactose © 2007 Paul Billiet ODWS
- 65. Adapting to the environment • E. coli can use either glucose, which is a monosaccharide, or lactose, which is a disaccharide • However, lactose needs to be hydrolysed (digested) first • So the bacterium prefers to use glucose when it can © 2007 Paul Billiet ODWS
- 66. Four situations are possible 1. When glucose is present and lactose is absent the E. coli does not produce β-galactosidase. 2. When glucose is present and lactose is present the E. coli does not produce β-galactosidase. 3. When glucose is absent and lactose is absent the E. coli does not produce β-galactosidase. 4. When glucose is absent and lactose is present the E. coli does produce β-galactosidase © 2007 Paul Billiet ODWS
- 67. Adapting to the environment • E. coli can use either glucose, which is a monosaccharide, or lactose, which is a disaccharide • However, lactose needs to be hydrolysed (digested) first • So the bacterium prefers to use glucose when it can © 2007 Paul Billiet ODWS
- 68. Four situations are possible 1. When glucose is present and lactose is absent the E. coli does not produce β-galactosidase. 2. When glucose is present and lactose is present the E. coli does not produce β-galactosidase. 3. When glucose is absent and lactose is absent the E. coli does not produce β-galactosidase. 4. When glucose is absent and lactose is present the E. coli does produce β-galactosidase © 2007 Paul Billiet ODWS
- 71. 1. When lactose is absent • A repressor protein is continuously synthesised. It sits on a sequence of DNA just in front of the lac operon, the Operator site • The repressor protein blocks the Promoter site where the RNA polymerase settles before it starts transcribing Regulator gene lac operon Operator site z y a DNA I O Repressor protein RNA polymerase Blocked © 2007 Paul Billiet ODWS
- 72. 2. When lactose is present • A small amount of a sugar allolactose is formed within the bacterial cell. This fits onto the repressor protein at another active site (allosteric site) • This causes the repressor protein to change its shape (a conformational change). It can no longer sit on the operator site. RNA polymerase can now reach its promoter site z y a DNA I O © 2007 Paul Billiet ODWS
- 73. 2. When lactose is present • A small amount of a sugar allolactose is formed within the bacterial cell. This fits onto the repressor protein at another active site (allosteric site) • This causes the repressor protein to change its shape (a conformational change). It can no longer sit on the operator site. RNA polymerase can now reach its promoter site Promotor site z y a DNA I O © 2007 Paul Billiet ODWS
- 74. 3. When both glucose and lactose are present • This explains how the lac operon is transcribed only when lactose is present. • BUT….. this does not explain why the operon is not transcribed when both glucose and lactose are present. © 2007 Paul Billiet ODWS
- 75. • When glucose and lactose are present RNA polymerase can sit on the promoter site but it is unstable and it keeps falling off Promotor site z y a DNA I O Repressor protein removed RNA polymerase
- 76. 4. When glucose is absent and lactose is present • Another protein is needed, an activator protein. This stabilises RNA polymerase. • The activator protein only works when glucose is absent • In this way E. coli only makes enzymes to metabolise other sugars in the absence of glucose Promotor site z y a DNA I O Transcription Activator protein steadies the RNA polymerase © 2007 Paul Billiet ODWS
- 77. Summary Carbohydrates Activator protein Repressor protein RNA polymerase lac Operon + GLUCOSE + LACTOSE Not bound to DNA Lifted off operator site Keeps falling off promoter site No transcription + GLUCOSE - LACTOSE Not bound to DNA Bound to operator site Blocked by the repressor No transcription - GLUCOSE - LACTOSE Bound to DNA Bound to operator site Blocked by the repressor No transcription - GLUCOSE + LACTOSE Bound to DNA Lifted off operator site Sits on the promoter site Transcription © 2007 Paul Billiet ODWS
- 79. NEGATIVE REGULATION X INDUCIBLE TRANSCRIPTION X REPRESSIBLE TRANSCRIPTION X
- 80. X NEGATIVE REGULATION INDUCIBLE TRANSCRIPTION X REPRESSOR INDUCER INACTIVE REPRESSOR REPRESSOR BINDING SITE OPERATOR The lac Operon
- 81. The lac Operon P O lacZ lacY lacA P O lacZ lacY lacA mRNA 5’ 3’ RIBSOSOME BINDING SITE Q: How many proteins are made?
- 82. The lac Operon P O lacZ lacY lacA P O lacZ lacY lacA mRNA 5’ 3’ Proteins b-galactosidase Permease Transacetylase
- 83. The lac Operon -b-galactosidase and permease encode proteins that metabolize lactose to give glucose. -In the absence of lactose in the medium, the genes are turned off. That is, no mRNA is transcribed and no proteins are made. -In the presence of lactose in the medium, the genes are turned on. That is, mRNA is transcribed and the proteins are made. X X REPRESSOR INDUCER INACTIVE REPRESSOR OPERATOR Q: Which molecule do you think is the inducer?
- 84. The lac Operon P O lacZ lacY lacA lac repressor X NO mRNA lac repressor -Protein that is encoded by the lacI gene. -The lacI gene has its own promoter. -The lac repressor binds to the operator and inhibits transcription of the lac operon.
- 85. The lac Operon In the presence of lactose in the medium, lactose binds to the repressor. The lactose-repressor complex is unable to bind to the operator. P O lacZ lacY lacA X NO mRNA lactose P O lacZ lacY lacA X NO mRNA
- 86. The lac Operon The RNA polymerase can now bind to the promoter and initiate transcription of the genes. The proteins made metabolize the lactose. P O lacZ lacY lacA mRNA
- 87. NEGATIVE REGULATION X INDUCIBLE TRANSCRIPTION X REPRESSIBLE TRANSCRIPTION X
- 90. NEGATIVE REGULATION REPRESSIBLE TRANSCRIPTION X Aporepressor Co-repressor Active repressor Operator THE trp OPERON
- 91. X Aporepressor Co-repressor Active repressor Operator THE trp OPERON -In the presence of tryptophan in the cell, the genes are turned off. That is, mRNA is not transcribed and the proteins are not made. -In the absence of tryptophan in the medium, the genes are turned on. That is, mRNA is transcribed and proteins are made. Q: Which molecule is the co-repressor?
- 93. Transcriptional Activators - Are proteins that activate transcription (No Dah!). - Bind to enhancers (in eukaryotes). - Their activity is modulated by environmental conditions. Enhancers (in eukaryotes) - Are specific DNA sequences that bind to transcriptional activators. - Can be found upstream or downstream of the promoter. - Do not need to be next to promoter.
- 94. GENRAL NOTES 1) Positive and negative regulation is used for all kinds of genes, polycistronic and monocistronic. It is used in all organisms. 2) Most genes have both positive and negative regulation (for example, lac operon). 3) When transcription is repressed, there is a very low level of mRNA made. That is called basal transcription.
- 95. E. coli changes gene expression to utilize different nutrient sources
- 96. Lac Operon (Negative Regulation) Catabolic Enzyme Repressor Inducer Operon
- 97. The lac repressor dissociates from the operator sequence upon IPTG (inducer) binding
- 98. Cis versus Trans
- 99. Cis versus Trans Dominant versus Recessive
- 100. Operator Mutations Reduce Repressor Binding
- 101. Fig. 7.12 The lac repressor tetramer binds two operators
- 102. Figs. 7.16 & 7.13 The lac operon is also regulated by glucose levels Low Glucose => High cAMP => activation of lac operon thru CAP binding site Glucose is Sweeter
- 103. cAMP is Needed for Activation Catabolite Repression
- 104. Figs. 7.17 & 7.19 CAP-cAMP binding creates DNA bending and activates lac operon transcription
- 106. Figs. 7.28 &7.31 The trp operon attenuator
- 107. Termination No termination Fig. 7.30 Alternative structures of the trp operon attenuator RNA
- 109. Reading: MVA pp. 760-767 and 1011-1018 Problem Ch 26: 9 Alberts: pp. 395-399
- 110. MVA Fig.21.1
- 111. MVA Fig.21.12
- 112. MVA Fig.21.13
- 113. MVA Fig.21.14
- 114. MVA Fig.21.15
- 115. The chromosomal order of genes in the trp operon of E. coli and the sequence of reactions catalyzed by the enzyme products of the trp structural genes. The products of genes trpD and trpE form a complex that catalyzes specific steps, as do the products of genes trpB and trpA. Tryptophan synthetase is a tetrameric enzyme formed by the products of trpB and trpA. It catalyzes a two-step process leading to the formation of tryptophan. (PRPP, phosphoribosyl pyrophosphate; CDRP, 1-(o- carboxyphenylamino)-1-deoxyribulose 5-phosphate.) (After S. Tanemura and R. H. Bauerle, Genetics 95, 1980, 545.)
- 119. MVA Fig. 26.33
- 120. Alberts Fig. 7-34
- 122. Originally, regulation of the trp operon was thought to occur solely through the repressor-operator system until deletion mutants located downstream of trpO were identified. These mutants displayed increased expression of the operon by six-fold which indicated the presence of an additional transcriptional control element. Why is repression not the only mode of regulation?
- 123. vidence: 1. Trp-tRNA synthetase mutants had regulatory anomalies. 2. Addition of trp to trp-starved cells not only shut down initiation of transcription but also inhibited transcription already in progress on the initial segment of the operon. 3. Mutants lacking a functional repressor could still respond to trp starvation by increasing transcription of trp mRNA. 4 . Deletion mutants in which both of the deletion termini were within the transcribed region of the operon had an unexpected six-fold increase in expression of the remaining genes in the operon. Obviously, repressor binding was unaffected.
- 124. 5. Within the population of mRNAs produced in vivo from the 5' end of the trp operon, RNAs corresponding to the first 140 bp (the leader sequence) of the operon were several times more abundant than those from more distal regions, therefore a transcription termination site was located before the structural genes. 6. Starving bacteria of trp reduced termination at this site (the trp attenuator). 7. Mutations altering trp-tRNA synthetase, tRNAtrp or a tRNA trp modifying enzyme were found to decrease transcription termination at the trp attenuator. What does this suggest about the mode of attenuation?
- 125. 8. Ribosome binding experiments with the 140 base transcript demonstrated that ribosomes protect a 20 base segment from nuclease attack. A potential AUG start codon is located in the center of this region. 9. A 14 residue peptide (the leader peptide) could be synthesized from this start codon and contained tandem trp residues near its C-terminus. 10. The trp leader ribosome binding site was shown to be an efficient site for the initiation of translation by fusing the leader to a structural gene and demonstrating synthesis of the fused polypeptide.
- 129. 11. Two classes of termination defective leader mutants have been isolated. One type terminates at less than normal frequency and has bp changes in the 3:4 bp region. In vivo, these mutants have a 2-4 fold increase in operon expression. 12. The second class of mutants have increased termination of the attenuator. These prevent the relief from termination that is associated with trp starvation. One of these mutants has an altered start codon for the leader peptide. Another has a G to A conversion at position 75, which would prevent 2:3 pairing and cause formation of a 3:4 termination structure.
- 130. Genetic analysis indicated that the new control element was located in trpL, a 162 nt region 30-60 nt upstream from trpE. When trp is scarce, the entire 6720 nt polycistronic trp, including trpL, is synthesized. As the trp concentration increases, the rate of trp transcription decreases as a result of the trp r epressor-corepressor's greater abundance. With increasing [trp], the mRNA synthesized consists more and more of a 140 nt segment corresponding to trpL sequences only. The availability of trp results in the premature termination of transcription of the operon.
- 131. MVA Fig. 26.35
- 133. MVA Fig. 26.36
- 135. MVA Fig.21.17
- 136. MVA Fig. 26.37
- 138. MVA Fig. 26.4
- 139. How a Terminator Loop is Formed
- 140. Lambda can Lyse bacterial Cells
- 141. Tight Regulation
- 142. Anti-terminator - A Friend in Disguise N
- 143. How to Start the Feedback Loop CI CI/CIII
- 144. Feedback Loop is Maintained Lamda lives Happily Thereafter… CI
- 145. Lysogeny versus Lysis CI wins - Cro wins - Lysogeny Lytic
- 146. What If Stock Market Turns South ? SOS RecA+ Lambda Protease
- 147. DNA-Protein
- 148. a-Helix Recognizes DNA Major Groove
- 149. DNase Footprinting Reveals Site of Binding
- 150. Amino-acid Side-chains Contact Functional Groups of Nucleotides
- 151. Major Groove - A Rich Source of Specificity
- 152. RNA DNA PROTEIN RNA
- 153. Stages of Transcription - Initiation
- 154. Stages of Transcription – Elongation and Termination
- 155. Bacterial Promoters • A promoter is where RNApoly binds • Template strand versus Coding strand • Upstream – in the 5’ direction on the coding strand • Consensus sequences
- 161. Initiation • Stayes put for a bit • Nucleoside triphospates (NTPs) are added one by one until 9 bp have been added, and then elongation can occur (i.e. RNApoly can move)
- 162. Elongation • Unwinding of DNA • Adding NTPs • About 12 bp of RNA are bonded to DNA as it goes • DNA is rewound behind • RNApoly moves along the template strand from 3’ to 5’, thus RNA elongates from the 5’ to 3’ end
- 163. Termination • The terminator is in the transcript, not the DNA • Forms a hairpin • Self- complementary • The hairpin structure is the signal for termination
- 169. Eukaryotic transcription • Three RNA polymerases • One for each major type of RNA – RNApoly I - makes pre-rRNA – RNApoly II - makes pre-mRNA – RNApolyIII - makes pre-tRNA • Each polymerase has a different promoter structure
- 170. RNApoly II promoter • Initiator sequence surrounding the start point • TATA box at about –25 bp • TATA+Initiator = core promoter • A transcription factor binds to the TATA box before RNAPoly II
- 172. Transcription factors A basal transcription factor is always required to allow RNApoly to bind to DNA For RNApoly II, TFIID binds to the TATA box. This is the basal transcription factor. More TFs bind to TFIID through protein-protein interactions to form the pre-initiation complex. Then RNApoly binds Many TFs may be involved
- 178. mRNA modifications • The primary transcript is modified • 5’ caps • poly(A) tails • splicing
- 179. mRNA Modifications: 5’methylated cap • A “backwards” 5’ cap of methylated guanine • Added during elongation • Functions – Flag for nuclear export – Protect against degradation – Binding site for the ribosome • Catalyzed by a "capping enzyme" that only associates with RNA poly II • Why might this be important?
- 180. mRNA modifications: polyadenylation • Transcripts are generally too long • poly(A) polymerase (PAP) finds a poly(A) signal (AAUAAA) that marks the end of the important stuff. • PAP cleaves the transcript. • PAP then adds a polyA tail to the newly cleaved mRNA • The poly-A tail: – helps protect the transcript from degradation – is necessary for full initiation of translation
- 182. mRNA degradation and poly(A) tails G XRN • De-adenylation promotes rapid degradation DCP
- 185. Exons usually represent protein domains
- 186. Splicing • Introns need to be removed from the primary transcript
- 187. Spliceosome • Splicing introns from mRNA occurs at short, conserved sequences called splice sites which specify the beginning and ends of introns. • GU on the 5' end and AG on the 3' end are 100% conserved. • Catalyzed by a structure called a spliceosome, composed of protein and RNA (snRNA – small nuclear RNA)
- 188. Splicing Reaction
- 189. rRNA modifications
- 190. Regulation of transcription • Gene “expression” and subsequent protein production is controlled primarily at the level of transcription. • That is, if you transcribe the gene, you will make the protein. • MODY is a defect in insulin gene regulation, not in transcription per se.
- 191. Example in bacteria Goal: Express the enzymes necessary to use lactose as an energy source only if lactose is present in the environment
- 192. Operon model • Operon: Several genes with related functions that are regulated together, because one piece of mRNA codes for several related proteins. • polycistronic mRNA,, mRNA coding for more than one polypeptide, is found only in prokaryotes
- 193. Operator • The operator is a sequence located between the promotor and the first structural gene • Proteins may bind to the operator and promote or inhibit transcription.
- 194. Operon model • An operon consists of: – Structural genes - code for the enzymes/proteins of interest. – Regulatory genes - control the expression of structural genes by expressing regulatory proteins
- 195. The lac operon • Repressors – bind to the operator and inhibit • Negative control of transciption, because binding of a protein to the operator turns transcription off
- 197. The lac operon • Substrate induction, or operon de- repression
- 198. Example in bacteria • The lac operon is involved with utilizing lactose as an energy source (a catabolic pathway). • Anabolic pathways are also controlled • Example: stop manufacturing the amino acid tryptophan if you have enough of it, either from synthesis or from the environment.
- 199. The trp operon • In the presence of tryptophan, the repressor for the trp operon binds to the operator. • Example of end product repression
- 200. Review… • In substrate induction, substrate (effector) binding to the repressor renders it unable to bind to the operator, and allows transcription to initiate. • In end-product repression, product (effector) binding to the repressor makes it bind. This prevents initiation of transcription. • These are both negative forms of regulation
- 201. • Glucose pathways are constitutively expressed • If glucose is present, turn down other pathways (like lactose catabolism) • If absent, only turn up those pathways for which there are substrates (like lactose) • Example: CRP in lac that promotes polymerase binding! Glucose = cAMP Positive control of the lac operon
- 202. Test yourself… • What would happen in the lac operon if… – There IS glucose, and there is NOT lactose? – There is NOT glucose, and there IS lactose? – There IS glucose, and there IS lactose?
- 203. A few more terms… • Regions in/around the promoter to which regulatory proteins may bind are called cis- acting elements. • Regulatory genes are often called trans-acting elements, because they can exist far away on the DNA – Their products, regulatory proteins, diffuse and bind to the cis-elements. – The gene for a repressor is one example.
- 204. Regulatory elements in eukaryotes • The basic principles that control transcription in bacteria also apply to eukaryotic organisms: – controlled by trans-acting proteins (transcription factors) binding to cis-acting DNA sequences • However, – eukaryotic cis-acting elements are often much further from the promoter they regulate, – Any given transcription factor may be involved as one of many TFs in the transcription of many different genes.
- 205. Complex control of eukaryotic genes Trans-acting element TFA TFA TFA Trans-acting element TFB TFB
- 206. Proximal control elements • The core promoter can typically turn on transcription only at a low rate. Add to these… • Proximal control elements – sequences <100 bases upstream of the core promoter to which transcription factors bind and improve the efficiency of the core promotor. • Transcription factors that bind outside the core promotor are called regulatory transcription factors.
- 207. • > 100 bp upstream or downstream • Enhancers – TFs that bind here are activators • Silencers – TFs that bind here are repressors Distal control elements
- 208. Steroid hormones • Steroid hormones pass through the cell membrane • Bind to and activate (or un-inhibit) a steroid receptor • The receptor moves to the nucleus • The receptor acts as a transcription factor
- 209. The genetic code: how do nucleotides specify 20 amino acids? 1. 4 different nucleotides (A, G, C, U) 2. Possible codes: • 1 letter code 4 AAs <20 • 2 letter code 4 x 4 = 16 AAs <20 • 3 letter code 4 x 4 x 4 = 64 AAs >>20 3. Three letter code with 64 possibilities for 20 amino acids suggests that the genetic code is degenerate (i.e., more than one codon specifies the same amino acid).
- 210. The genetic code is a triplet code A set of 3 consecutive nucleotides make a codon in mRNA code, which corresponds to one amino acid in a polypeptide chain. 1. 1960s: Francis Crick et al. 2. Studied frameshift mutations in bacteriophage T4 (& E. coli), induced by the mutagen proflavin. 3. Proflavin caused the insertion/deletion (indels) of a base pair in the DNA. 4. Two ways to identify mutant T4: 1. Growth with E. coli B: • r+(wild type) turbid plaques • rII (mutant) clear plaques 2. Growth with E. coli K12 (): • r+ (wild type) growth • rII (mutant) no growth
- 211. 1. Discovered that frameshift mutations (insertion or deletion) resulted in a different sequence of amino acids. 2. Also discovered that r+ mutants treated with proflavin could be restored to the wild type (revertants). Fig. 6.5
- 212. 3. Combination of three r+ mutants routinely yielded revertants, unlike other multiple combinations. Fig. 6.6 - Three nearby insertions (+) restore the reading frame, giving normal or near-normal function.
- 213. How was the genetic code deciphered? 1. Cell-free, protein synthesizing machinery isolated from E. coli. (ribosomes, tRNAs, protein factors, radio-labeled amino acids). Synthetic mRNA containing only one type of base: UUU = Phe, CCC = Pro, AAA = Lys, GGG = ? (unstable) 2. Synthetic copolymers (CCC, CCA, CAC, ACC, CAA, ACA, AAC, AAA) composed of two different bases: Pro, Lys (already defined) + Asp, Glu, His, & Thr Proportion (%AC) varied to determine exactly which codon specified which amino acid. 3. Synthetic polynucleotides of known composition: UCU CUC UCU CUC Ser Leu Ser Leu 1968: Robert Holley (Cornell), H. G. Khorana (Wisconsin-Madison), and Marshall Nirenberg (NIH).
- 214. How was the genetic code deciphered (cont.): 4. Ribosome binding assays of Nirenberg and Leder (1964) (ribosomes, tRNAs charged w/AAs, RNA trinucleotides). Protein synthesis does not occur. Only one type of charged tRNA will bind to the tri-nucleotide. mRNA UUU codon tRNA AAA (with Phe) anti-codon mRNA UCU codon tRNA AGU (with Ser) anti-codon mRNA CUC codon tRNA GAG (with Leu) anti-codon Identified 50 codons using this method. 5. Combination of many different methods eventually identified 61 codons, the other 3 do not specify amino acids (stop-codons).
- 215. Fig. 6.7
- 216. Characteristics of the genetic code (written as in mRNA, 5’ to 3’): 1. Code is triplet. Each 3 codon in mRNA specifies 1 amino acid. 2. Code is comma free. mRNA is read continuously, 3 bases at a time without skipping bases (not always true, translational frameshifting is known to occur). 3. Code is non-overlapping. Each nucleotide is part of only one codon and is read only once. 4. Code is almost universal. Most codons have the same meaning in different organisms (e.g., not true for mitochondria of mammals). 5. Code is degenerate. 18 of 20 amino acids are coded by more than one codon. Met and Trp are the only exceptions. Many amino acids are four-fold degenerate at the third position. 6. Code has start and stop signals. ATG codes for Met and is the usual start signal. TAA, TAG, and TGA are stop codons and specify the the end of translation of a polypeptide. 7. Wobble occurs in the tRNA anti-codon. 3rd base is less constrained and pairs less specifically.
- 217. Wobble hypothesis: Proposed by Francis Crick in 1966. Occurs at 3’ end of codon/5’ end of anti-codon. Result of arrangement of H-bonds of base pairs at the 3rd pos. Degeneracy of the code is such that wobble always results in translation of the same amino acid. Complete set of codons can be read by fewer than 61 tRNAs. 5’ anti-codon 3’ codon G pairs with U or C C pairs with G A pairs with U U pairs with A or G I (Inosine) pairs with A, U, or C I = post-transcription modified purine Fig. 6.8
- 218. TTT TCT TAT TGT TTC TCC TAC TGC TTA TCA TAA TGA TTG TCG TAG TGG CTT CCT CAT CGT CTC CCC CAC CGC CTA CCA CAA CGA CTG CCG CAG CGG ATT ACT AAT AGT ATC ACC AAC AGC ATA ACA AAA AGA ATG ACG AAG AGG GTT GCT GAT GGT GTC GCC GAC GGC GTA GCA GAA GGA GTG GCG GAG GGG
- 221. Evolution of the genetic code: Each codon possesses an inherent set of possible 1-step amino acid changes precluding all others. As a result, some codons are inherently conservative by nature, whereas others are more radical. Phe, Leu, Ile, Met, Val (16 codons with T at 2nd pos.) possess 104 possible evolutionary pathways. Only 12 (11.5%) result in moderately or radically disimilar amino acid changes Most changes are nearly neutral because they results in substitution of similar amino acids. DNA sequences with different codons compositions have different properties, and may evolve on different evolutionary trajectories with different rates of substitution.
- 222. Evolution of the genetic code (cont.): On average, similar codons specify similar amino acids, such that single base changes result in small chemical changes to polypeptides. For example, single base changes in the existing code have a smaller average effect on polarity of amino acids (hydropathy/hydrophily) than all but 0.02% of randomly generated genetic codes with the same level of degeneracy (Haig and Hurst 1991, J. Mol. Evol. 33:412-417). The code has evolved to minimize the severe deleterious effects of substituting hydrophilic for hydrophobic amino acids and vice versa (this also is true for other properties). This is a good thing!!!
- 223. Translation-protein synthesis (Overview): 1. Protein synthesis occurs on ribosomes. 2. mRNA is translated 5’ to 3’. 3. Protein is synthesized N-terminus to C-terminus. 4. Amino acids bound to tRNAs are transported to the ribosome. Facilitated by: Specific binding of amino acids to their tRNAs. Complementary base-pairing between the mRNA codon and the tRNA anti- codon. mRNA recognizes the tRNA anti-codon (not the amino acid).
- 224. Translation - 4 main steps 1. Charging of tRNA 2. Initiation 3. Elongation (3 steps) 1. Binding of the aminoacyl tRNA to the ribosome. 1. Formation of the peptide bond. 1. Translocation of the ribosome to the next codon. 4. Termination
- 225. Step 1-Charging of tRNA (aminoacylation) 1. Amino acids are attached to tRNAs by aminoacyl-tRNA synthetase and Produces a charged tRNA (aminoacyl-tRNA). 2. Uses energy derived from ATP hydrolysis.(ATP=AMP+PPi) 3. 20 different aminoacyl-tRNA synthetases (one for each AA). 4. tRNAs possess enzyme-specific recognition sites for aminoacyl-tRNA synthetase 5. Sequence of events: 1. ATP and amino acid bind to aminoacyl-tRNA synthetase, to form aminoacyl- AMP + PPi. 2. tRNA binds to aminoacyl-AMP. 3. Amino acid transfers to tRNA, displacing AMP. 4. Amino acid always is attached to adenine on 3’ end of tRNA by its carboxyl group forming aminoacyl-tRNA.
- 227. Step 2-Initiation-requirements: 1. mRNA 2. Ribosome 3. Initiator tRNA (fMet tRNA in prokaryotes) 4. 3 Initiation factors (IF1, IF2, IF3) 5. Mg2+ 6. GTP (guanosine triphosphate)
- 228. Step 2-Initiation-steps (e.g., prokaryotes): 1. 30S ribosome subunit + IFs/GTP bind to AUG start codon and Shine-Dalgarno sequence composed of 8-12 purine-rich nucleotides upstream (e.g., AGGAGG). 2. Shine-Dalgarno sequence is complementary to 3’ 16S rRNA. 3. Initiator tRNA (fMet tRNA) binds AUG (with 30S subunit). All new prokaryote proteins begin with fMet (later removed). fMet = formylmethionine (Met modified by transformylase; AUG at all other codon positions simply codes for Met) mRNA 5’-AUG-3’start codon tRNA 3’-UAC-5’ anti-codon 4. IF3 is removed and recycled. 5. IF1 & IF2 are released and GTP is hydrolysed, catalyzing the binding of 50S rRNA subunit. 6. Results in a 70S initiation complex (mRNA, 70S, fMet-tRNA)
- 229. See 6.15
- 230. Step 2-Initiation, differences between prokaryotes and euakaryotes: 1. Initiator Met is not modified in eukaryotes (but eukaryotes possess initiator tRNAs). 2. No Shine-Dalgarno sequence; but rather initiation factor (IF-4F) binds to the 5’-cap on the mature mRNA. 3. Eukaryote AUG codon is embedded in a short initiation sequence called the Kozak sequence. 4. Eukaryote poly-A tail stimulates translation by interacting with the 5’-cap/IF-4F, forming an mRNA circle; this is facilitated by poly-A binding protein (PABP).
- 231. Step 3-Elongation of a polypeptide: 1. Binding of the aminoacyl tRNA (charged tRNA) to the ribosome. 2. Formation of the peptide bond. 3. Translocation of the ribosome to the next codon.
- 232. 3-1. Binding of the aminoacyl tRNA to the ribosome. • Ribosomes have two sites, P site (5’) and A site (3’) relative to the mRNA. • Synthesis begins with fMet (prokaryotes) in the P site, and aa-tRNA hydrogen bonded to the AUG initiation codon. • Next codon to be translated (downstream) is in the A site. • Incoming aminoacyl-tRNA (aa-tRNA) bound to elongation factor EF-Tu + GTP binds to the A site. • Hydrolysis of GTP releases EF-Tu, which is recycled. • Another elongation factor, EF-Ts, removes GDP, and binds another EF-Tu + GTP to the next aa-tRNA. • Cycle repeats after peptide bond and translocation.
- 233. Fig. 6.17
- 234. 3-2. Formation of the peptide bond. • Two aminoacyl-tRNAs positioned in the ribosome, one in the P site (5’) and another in the A site (3’). • Bond is cleaved between amino acid and tRNA in the P site. • Peptidyl transferase (catalytic RNA molecule - ribozyme) forms a peptide bond between the free amino acid in the P site and aminoacyl-tRNA in the A site. • tRNA in the A site now has the growing polypeptide attached to it (peptidyl- tRNA). Fig. 6.18
- 235. 3-3. Translocation of the ribosome to the next codon. • Final step of the elongation cycle. • Ribosome advances one codon on the mRNA using EF-G (prokaryotes) or EF-2 (eukaryotes) and GTP. • Binding of a charged tRNA in A site (3’) is blocked. • Uncharged tRNA in P site (5’) is released. • Peptidyl tRNA moves from A site to the P site. • Vacant A site now contains a new codon. • Charged tRNA anti-codon binds the A site, and the process is repeated until a stop codon is encountered. • Numbers and types of EFs differ between prokaryotes and eukaryotes. • 8-10 ribosomes (polyribosome) simultaneously translate mRNA.
- 236. Fig. 6.17
- 237. Fig. 6.19
- 238. Step 4-Termination of translation: 1. Signaled by a stop codon (UAA, UAG, UGA). 2. Stop codons have no corresponding tRNA. 3. Release factors (RFs) bind to stop codon and assist the ribosome in terminating translation. 1. RF1 recognizes UAA and UAG 2. RF2 recognizes UAA and UGA 3. RF3 stimulates termination 4. 4 termination events are triggered by release factors: 1. Peptidyl transferase (same enzyme that forms peptide bond) releases polypeptide from the P site. 2. tRNA is released. 3. Ribosomal subunits and RF separate from mRNA. 4. fMet or Met usually is cleaved from the polypeptide.
- 239. See Fig. 6.20