
Recomendados
Mais conteúdo relacionado
Mais procurados
Mais procurados (20)
Destaque
Destaque (10)
Semelhante a Cell Bio 1
Semelhante a Cell Bio 1 (20)
Mais de Mark Botirius
Mais de Mark Botirius (6)
Cell Bio 1
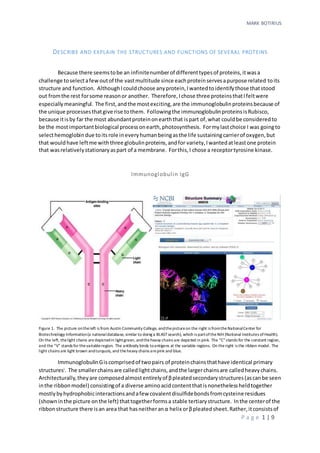
- 1. MARK BOTIRIUS P a g e 1 | 9 DESCRIBE AND EXPLAIN THE STRUCTURES AND FUNCTIONS OF SEVERAL PROTEINS Because there seemstobe an infinitenumberof differenttypesof proteins,itwasa challenge toselectafewoutof the vastmultitude since eachproteinservesapurpose related toits structure and function. AlthoughIcouldchoose anyprotein,Iwantedtoidentifythose thatstood out fromthe rest forsome reasonor another. Therefore,Ichose three proteinsthatIfeltwere especiallymeaningful. The first,andthe mostexciting,are the immunoglobulinproteinsbecause of the unique processesthatgive rise tothem. Followingthe immunoglobulinproteinsisRubisco, because itisby far the most abundantproteinonearththat ispart of,what couldbe consideredto be the mostimportantbiological processonearth,photosynthesis. Formylastchoice I was goingto selecthemoglobindue toitsrole ineveryhumanbeingasthe life sustainingcarrierof oxygen,but that wouldhave leftme withthree globulinproteins,andfor variety,Iwantedatleastone protein that wasrelativelystationaryaspart of a membrane. Forthis,I chose a receptortyrosine kinase. Immunoglobulin IgG Figure 1. The picture ontheleft is from Austin Community College, andthepictureon the right is fromtheNationalCenter for Biotechnology Information(a nationaldatabase, similar to doing a BLASTsearch), which is partoftheNIH (National Institutes ofHealth). On the left, thelight chains aredepictedin lightgreen, andtheheavy chains are depicted in pink. The “C”stands for the constant region, and the “V” stands for thevariableregion. The antibody binds toantigens at the variable regions. On theright is the ribbon model. The light chains are light brown andturquois, and theheavy chains arepink and blue. ImmunoglobulinGiscomprisedof twopairs of proteinchainsthathave identical primary structuresi . The smallerchainsare calledlightchains,andthe largerchainsare calledheavychains. Architecturally,theyare composedalmostentirelyof βpleatedsecondarystructures(ascanbe seen inthe ribbonmodel) consistingof a diverse aminoacidcontentthatisnonethelessheldtogether mostlybyhydrophobicinteractionsandafew covalentdisulfidebondsfromcysteine residues (showninthe picture onthe left) thattogetherformsa stable tertiarystructure. Inthe centerof the ribbonstructure there isan area that hasneitheranα helix orβpleatedsheet.Rather,itconsistsof
- 2. MARK BOTIRIUS P a g e 2 | 9 a flexible hinge,whichgivesthe moleculeelasticitythatservestoaugmentitsabilitytobindtoan antigen. Notice thatthe β pleatedsheetsof boththe heavyandlightchainsinthe ribbonmodel are connectedbysingle strandedloops. The loopslocatedatthe top rightand topleft of the molecule comprise the areasinthe variable regionswhere the antibodybindstoanantigen. Whenwe considerthe variousstructural aspectsof thismolecule,itisclearthat thismolecule isstructurally adaptedto functionasa bindingmolecule. The β pleatedsheetsgive itthe necessarystrengthand stability,andyetthe loopingsingle strandsgive itthe neededflexibilitytobindstronglytoan antigen. Flexibilityisalsofoundatthe centerof the molecule atthe hinge. What isso amazingabout thisantibodymolecule (andalsoaboutall antibodymoleculesin general) isthatif youpicture the molecule withoutitsvariable lightandheavyregionsonthe ends,it isclear that itis an excellentplatformforamolecule whose functionisto bindavarietyof other molecules. AsIhave just described,itisbothstructurallystable andyetflexible. Itissimilartohow a multi-tipscrewdriverhandle isanexcellentplatformtoturnvariousscrew types. If youneedto turn a standardscrew,you popon a standardtip. If you needtoturn a Phillip’sscrew,youpopona Phillip’shead. Incredibly, thisissortof whathappensinour immune systems.The difference isthat our immune systemsaren’tmakingtipstofita particularscrew,they make as manydifferentkinds of tipsas possible,inthe eventtheyrunintoa“pathogenscrew”that fitstheirparticulartip. The genesthatencode the variable regionsatthe endsof the “Y” onthe antibodymolecule are composedof manysegments. These segmentsare differentforlightandheavychains;however,the principle isthe same. Forexample,the lightvariablechainsegmentsare comprisedof two categories:V andJ. There are several different“V”segments,andseveral different“J”segments. Onlyone of each isbroughttogetherina developingBcell tocreate (literally) the gene forthe variable region. Inotherwords,the gene thatcodesforthe variable regioniscreatedinthe Bcell throughgenetic recombination. Itturnsout,naturehasbeen in thegenetic engineering business long beforemankind waseven a twinklein theevolutionary eye! In thisway,the immune system can theoreticallycreate anantibodywithinagivenpopulationforalmostanypathogeninexistence, by creatingdifferent“tips”(the variable regionsonthe ends) andattachingthemtoa standard “platform”(the constantregions). V1 V2 V3 J1 J2 J3 J4 C Variable Region ConstantRegion Figure 2. The top row represents the region of DNA that contains the various V,J, and C segments. Through DNA recombination,one of each of the V, J, and C segments is broughttogether to form a contiguous gene represented by the bottom row. Because each gene is different, they each code for a di fferent variableregion, which can bind to a different antigen. Rubisco V3 J2 C
- 3. MARK BOTIRIUS P a g e 3 | 9 Because RuBisCo(ribulose 1, 5 bisphosphate carboxylase/ oxygenase)isthe onlyknown proteinthatcan fix CO2 in higherplants,itisarguablythe mostabundantproteinon earth. Despite itsimportant role inthe biochemistryof life, it isa rather slow and inefficientenzyme. On average,RubisCocanonly catalyze 3 to ten molecules each second (Cambridge CAPP, 2016). One reasonisbecause RuBisCo,asits unabbreviated name indicates,isbotha carboxylase andanoxygenase. Althoughitwill preferentially bindCO2,it doessoonly slightly,andsointhe presence of oxygenitnotonlyattaches carbon to RuBP,it will attach oxygenaswell,whichisa costlymove forthe plantbiochemically. Toprocessthe oxidizedRuBPmolecule(called 2- phosphoglycolate)the plantendsuplosingCO2. The primarystructure of thisproteinisalsohighlyvaried,andthe secondarystructure,as depictedinfigure 3,consistsof bothα helicesandβpleatedsheetsinadditiontoloops. Ananalysis of the secondarystructure revealsthatthe α helicesandβpleatedsheetslocatedinthe centerare hydrophobic,while the loopslocatedalongthe peripheryandinthe centerhole are mostly hydrophilic. ThisallowsRuBPaccesstothe active siteswhichare locatedonthe large subunitsof the molecule towardsthe center. These active sitesare constructedof β barrelssurroundedbyα helices. The βbarrels are the catalyticcenters of the active site that holda Mg2+ ionin the catalyticpocketby interactingwiththe polarchargedamino acidshistidine andlysine. Thisisa good example of the relationshipbetweenstructure andfunction. The creationof a six carbon intermediate sugarfromRuBPisaccomplishedviaa chargedmetal ion. Thisis the reasonthat we findchargedpolar aminoacidsin the catalyticsite. WhenCO2 entersthe active site, it attachesto the lysine residue,whichdestabilizesthe metal ion that resultsin a conformational change thatbringsthe RuBP molecule andCO2 moleculetogether,catalyzingthe reactionthat formsa six carbon sugar. (PNAS,2017) (Goodsell,2017) Tyrosine Kinase Receptor Figure 3. This is a ribbon model for the RuBisCo protein found in a spinach plant. Itis composed of 8 largesubunits and 8 small subunits. Figure 4. Ribulose 1,5 bisphosphate carboxylase/oxygenase active site
- 4. MARK BOTIRIUS P a g e 4 | 9 Lastly,I chose a receptorproteinbecause it demonstratesnicelythe connectionbetween structure and function;since there isa distinctdifference betweenthosestructures, for example,thatare inside the membrane of a cell incontrast to those that are stickingout fromthe ends. Tyrosine kinase receptorsare so namedbecause theyare a signaling molecule that,whenboundtotheirparticular ligand,phosphorylate (enzymesthat phosphorylate othermoleculesare called kinases) theirtyrosine residues. RTK’sare amphipathicintegral membrane proteinsthat consistof three domains:anouterdomain that protrudesoutside of the cell andserves to bindligands,amiddle domainthatis situatedinside the cellmembrane,andan innerdomainthat protrudesintothe inside of the cell that servesto deliverthe signal received fromthe ligand. Thisisan excellentexampleof the connectionbetween structure and function. The outer endsof the molecule are incontact withthe aqueous environmentsthatexist bothwithinandwithout the cell. The middle section,hasa primary structure made up entirely of non-polaraminoacids, that have adoptedanα helical secondary structure. Thisnonpolar middle sectionisindirect contact withthe non-polar phospholipidtailsthat make up the intermembranespace. This,alongwithan α helical secondarystructure,securesthe proteininthe membrane. If you lookcarefullyatthe actual canonical sequence,youwillnotice thatthe tyrosine residuesare missing. Thatisbecause thisisthe actual sequence fromthe papershowninthe NCBI Figure 5. An NCBI rendered ribbonmodel of a tyrosine kinase receptor Figure 6 This is a tubular modelof the protein depicted in figure 5. The canonical sequence shown is the actual sequence ofthe protein, andthe highlighted areas depict the hydrophobic regionof the molecule to clearlyshow the connection betweenthe structure and function ofthe amino acids.
- 5. MARK BOTIRIUS P a g e 5 | 9 picture infigure five,whichfocusedspecificallyonthe transmembrane region. The shortened sequence alsomade iteasierforme toput the aminoacid sequence inthe text,sothatI couldmore clearlydemonstrate the hydrophobicstructure –functionconnection. The overall receptorfunction of the molecule ismore clearlyshowninthe followingfigure. As can be seeninthe figure above,whenaligandbindstoreceptordomainsof the proteins, a conformational change bringsthe totwodimerstogether. Asa result,the twodimersactually phosphorylate eachother,activatingthem. The activatedkinasesthenphosphorylate another molecule,which,whenactivatedphosphorylatesyetanothermolecule downthe cascade. One possible resultof the conclusionof the cascade isthe activationof some transcriptionfactorthat influencesgeneticexpression. Itisno accidentthatthe aminoacidtyrosine isfoundinside the cell. Tyrosine, serine,andthreonineare almostexclusivelythe active residuesinthe phosphorylation activitiesof proteins. Why?Because these are the onlyaminoacidswithanOH group as part of theirside chains,andthe hydroxyl groupiswhere the phosphatesare added(orremoved). Likewise,manyof the extracellulardomainsof kinaseshave polarand/orchargedresidues. The reasonisbecause theyoftenneedtobe able tobindwithligandmoleculeswithanaffinitythatis greaterthan that of unchargedor nonpolarmolecules. EXPLAIN AND SHOW HOW THE REACTIONS OF THE CENTRAL METABOLISM (GLYCOLYSIS, TCA CYCLE, OXIDATIVE PHOSPHORYLATION) INTERACT WITH ONE ANOTHER Figure 7 The two blue structures are the twoRTKdomains pictured as pinkandblue infigure 5. Thispicture is from “Membranereceptors.com
- 6. MARK BOTIRIUS P a g e 6 | 9 Aftercareful consideration,Ihave concludedthatglycolysisisthe mostlikelyplace tobegin, because itscarbonstartingmaterial (glucose) isnotprovidedbythe othertwopathways,andof the three,itappearsto have the fewestmoleculesfedintoitfromthe others. My biggestconcern,is that my answeriscohesive,organized,andclear. To ensure this,Ihave decidedthatthe bestwayis to addresseachreactionseparately,stepbystep,andasktwoquestions.1. What, if anything,did the othertwo reactionscontribute tothisstep? 2. What, if anything,doesthisstepcontribute to the othertwo? Glycolysis 1. Glycolysisbeginswithglucose asitsstartingmaterial. Glucose isphosphorylatedbythe enzyme hexokinase,usingamolecule of ATPinthe process. a. What, if anything,didthe othertworeactionscontribute tothisstep? The glucose came from sourcesoutside the three reactionsunderconsideration,sotheydidnot contribute the glucose. The ATP,theoretically,couldhave come fromanyof the three,since all three produce ATP. Since glycolysisalsoproducesATP,Iwouldsaythat mostlikely,the ATPcame fromglycolysisitself. b. What,if anything,doesthisstepcontributetothe othertwo? The product fromthisreaction(Glucose 6 phosphate) isnotusedineitherthe TCA or ETC reactions. Sonothingispassedonto the other two. Since the ETC needsADPto make ATP,it is possible thatthe ADPiscontributedtooxidativephosphorylation. 2. In the nextstepinglycolysis,fructose6phosphate isproducedfromglucose 6phosphate via the enzyme phosphoglucoseisomerase. The startingmaterial (glucose6phosphate) came fromthe previousstep,andthe productisnotusedin eitherthe TCA or ETC reactions. Therefore,the answertobothquestionsisnone. 3. Next,fructose 6 phosphate isphosphorylatedbythe enzyme phosphofructokinaseto produce fructose 1,6 bisphosphate. The answerstobothquestionsare the same as instep one,that is,none. 4&5. Next,fructose 1,6 bisphosphateissplitintotwothree carbonmolecules. One is glyceraldehyde 3-phospate,andthe otherisan intermediate metabolite,dihydroxyacetone phosphate. Dihydroxyacetone phosphate isthenisomerizedintoglyceraldehyde3- phosphate. Sooverall,thisreactionproduces2moleculesof G3P. It shouldbe noted,that the productsof thisreaction(G3P) are intermediatesinotherreactionsnotunder considerationhere,suchasthe Calvincycle. However,the answerstobothquestionsatthis pointisstill none. 6. In thisstep,bothmoleculesof G3Pare oxidizedbyNAD+ reducingtwomoleculesof NAD+ to NADH+ H+ . The productis 1,3 bisphosphoglycerate. a. What, if anything,didthe othertworeactionscontribute tothisstep? In the presence of oxygen,onlythe ETCproducesNAD+ . The othertwo reactions,glycolysis and TCA,consume it. In the absence of oxygen,however,the endproductof glycolysis,pyruvate,
- 7. MARK BOTIRIUS P a g e 7 | 9 can be reducedtoproduce NAD+ . Therefore,the NAD+ couldhave come fromthe ETC or fermentation. b. What,if anything,doesthisstepcontributetothe othertwo? The NADH moleculescarry electronsthatare usedinthe ETC reactionsto drive protonsinto the intermembrane space of the mitochondriontoproduce aprotongradient. It ispossible,the H+ moleculesalsohelpcontributetothe gradient. 7. In thisstep3-phosphoglycerate isproducedfrom1,3-bisphosphoglycerate producingone twomoleculesof ATPfromADP(remember,everythingfromsteps4 & 5 involve twomolecules). I considerthese moleculesof ATPtobe whatwas neededtobringthe glycolysisbalance to0. Therefore, the answertoquestionsaandb are none. 8, 9, & 10. None of the intermediate productsinthesestepsare usedinthe othertworeactions. The onlythingworthnotingis thattwo more moleculesof ATPare producedfromADP. Since ADP resultsfrom the utilizationof energy,itisprettymuchubiquitous,andtherefore there isnoreal needtoconnectit to any particularreaction. Lastly,the endproductof glycolysis,pyruvate,isused inthe TCA reaction. The TCA Cycle A careful lookatthe TCA cycle revealsthatit isessentiallylittle more thananelectroncarrier producingreaction. The onlyexceptionisthatitusesone GDP molecule toproduce GTP. It neither producesor useseitherATPorADP. Furthermore,it’sintermediatemetabolitesare notusedby glycolysisorthe ETC. Therefore,we needtoonlysummarizeitsusesof NAD+ ,FADand GDP to account forits relationshiptoglycolysisandthe ETC. The TCA producesNADHfrom NAD+ inthe followingreactions: 1. Oxidizingpyruvate toproduce Acetyl CoA 2. Oxidizingisocitrate toproduce α ketoglutarate 3. Oxidizingα ketoglutarate toproduce succinyl CoA 4. Oxidizingmalate toproduce oxaloacetate All of the NAD+ for these reactionswassuppliedbythe ETC. Glycolysis(fermentation notwithstanding) andthe TCA reactionsdonotproduce NAD+ . The NADHproducedisusedbythe ETC. The TCA cycle alsoproducesthe electroncarrierFADH2 from FADwhenitoxidizessuccinate to produce fumarate. Again,the FADissuppliedbythe ETC,and the FADH2 isusedbythe ETC. Lastly,a molecule of GDPisphosphorylatedwhenamolecule of succinate isproducedfromsuccynil CoA. This isthe onlyplace that we findGDP,and it isusedextensivelythroughoutthe cell,notjust inthisreaction. Oxidative Phosphorylation
- 8. MARK BOTIRIUS P a g e 8 | 9 The oxidative phosphorylationcycle consistsof aseriesof membrane boundelectron carriersthat are arrangedin orderfromthe one withthe greatestredox potential tothe one with the least. NADHand FADH2 give theirelectronsto the carrierswiththe greatestpotential thatcan accept theirelectrons. These carriersthenuse the energytoshuttleprotonsacrossthe membrane and thenpassthe electronstothe nextcarrier,whichdoesthe same. Finally,atthe endof the chain,the electronsare passedtoan oxygen. Therefore,the ETCusesthe NADH andFADH2 producedbyglycolysisandthe TCA reactionstomake NAD+ and FAD that isusedby glycolysisand the TCA. The protonsare usedby ATPsynthase tomake ATP fromADP. ADPis produced throughoutthe cell. The followingfiguresummarizestheseinteractions. Glycolysis The Citrus Acid Cycle Oxidative Phosphorylation NAD+ FAD ATP NADH FADH2 ATPGDP GTP NAD+ from fermentation
- 9. MARK BOTIRIUS P a g e 9 | 9 References (2016, September25).RetrievedfromCambridge CAPP: https://cambridgecapp.wordpress.com/improving-photosynthesis/rubisco/ (2017, September25).RetrievedfromPNAS:http://www.pnas.org/content/109/46/18785.full#F4 Goodsell,D.(2017, September25).RetrievedfromPDB101:http://pdb101.rcsb.org/motm/11 i For example, the canonical sequencefor the heavy chain pictured in the ribbon model is: 1 qvqlvqsgaevkkpgasvkv scqasgyrfsnfvihwvrqa pgqrfewmgw inpyngnkef 61 sakfqdrvtf tadtsantay melrslrsad tavyycarvgpyswddspqd nyymdvwgkg 121 ttvivssastkgpsvfplap sskstsggta algclvkdyf pepvtvswns galtsgvhtf 181 pavlqssgly slssvvtvps sslgtqtyic nvnhkpsntk vdkkaepksc dkthtcppcp 241 apellggpsv flfppkpkdtlmisrtpevtcvvvdvshed pevkfnwyvd gvevhnaktk 301 preeqynsty rvvsvltvlh qdwlngkeyk ckvsnkalpa piektiskak gqprepqvyt 361 lppsrdeltk nqvsltclvk gfypsdiavewesngqpenn ykttppvlds dgsfflyskl 421 tvdksrwqqg nvfscsvmhealhnhytqks lslspgk