2. The Nobel Prize
in Physiology or
Medicine 1983
was awarded to
Barbara
McClintock "for
her discovery of
mobile genetic
elements".
Barbara McClintock
4. Transposons and Insertional
Mutations
Transposons: Mobile Genetic
Elements
Transposon
Transposon Transgenesis
Barbara McClintock
Transposon
Insertional
Transposon Mutagenesis
chromosome 基 Gene 因
Mutant Gene Tagged
4
5. Advantages of Insertional Mutations
can produce easily tractable
mutations
can produce large number of
mutants at low cost and high speed
5
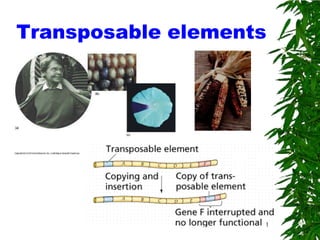
6. What are Transposons?
Transposable element (transposon): a sequence of DNA that is com
from place to place within a genome
Transposition of DNA on chromosome 9 of maize explains mottl
6
Some definitions and figures from Lisch 2009: Annu. Rev. Plant Biol. 2009.60:43-66.
7. What are Transposons?
Transposable element (transposon): a sequence of DNA that is com
from place to place within a genome
(1) At the beginning of kernel development, the Ds transposon is
inserted into the colored (C) gene, resulting in colorless tissue. (2) Ds
transposition early in kernel development restores the C gene, giving
rise to a large colored sector. (3) Transposition later in kernel
development results in smaller sectors. 7
Learn more at: weedtowonder.org/jumpingGenes.html
8. What are Transposons?
Transposable element (transposon): a sequence of DNA that is com
from place to place within a genome
“Cut & Paste”
“Copy & Paste” 8
9. What are Transposons?
• Plant genomes contain multiple transposon families.
• Each contains autonomous and non-autonomous elements.
• Class I transposons do not move, but are being copied.
• Class II transposons move, but can undergo copying, too (if
transposing during DNA replication)
Autonomous
element
Gene(
s)
Nonautonomous
elements
9
10. What are Transposons?
Transposons make up the major content of eukaryotic genomes
• ~50% of the genomes of human, chimp, mouse, ape
• ~75% of the maize genome
• ~85% of the barley genome
• ~98% of the iris genome
Iris brevicaulis Iris fulva
10
13. How do organisms live with TEs?
• Most TEs are broken (cannot tranpose; “fossils”).
• Active TEs evolved to insert into “safe-havens.”
• Host regulates TE movement.
• TEs can provide advantages.
13
14. Ping/mPing
mPing: MITEs are being amplified to
high copy numbers
MITE (Multi-insertional TE)
Deletion-derivative of Ping
Requires Ping transposase to jump 14
15. mPing copy number in O.japonica
OVER 1000 mPing copies
mPing
Japonica strains
Over 1000 copies of mPing in 4 related strains….
15
Naito et al PNAS (2006))
Takatoshi Tanisaka lab (Kyoto
16. Genomic distribution of mPing insertions
• predominantly in genic regions in euchromatin
• even inserts in heterochromatin are in genes
• where does mPing insert in and around genes?
16
17. Genic distribution of mPing insertions
12
shared
(n=926)
10
unshared
8 (n=736)
expect.
(%)
6
4
2
0
5' TR
U exon i
ntron 3' TR
U
UTR Exon UTR
17
mPing insertions rare in coding-exons
18. TEs can alter gene expression
Os02g0135500 (-41)
2.5
NB
EG4 (mPing+)
2 A123 (mPing+)
A157
1.5
1
0.5
0
control cold salt dry
18
mPing found to confer cold and salt inducibility
19. TEs can alter gene expression
Can this have phenotypic consequences?
Nipponbare EG4
19
EG4 is salt tolerant
20. Rapid mPing amplification (burst)
• Massive amplification largely benign
• Subtle impact on the expression of many genes
• Produces stress-inducible networks (cold, salt, others?)
• Generates dominant alleles
Naito et al, Nature, 2009
20
21. TEs as tools of evolutionary change
• TEs usually inactive.
• “Stress” conditions may activate TEs.
• Active TEs increase mutation frequency.
• Most mutations caused by TEs neutral or harmful.
• A rare TE-induced mutation (or rearrangement) may be adaptive.
Transposable elements can shake up otherwise conservative
genomes and generate new genetic diversity.
21
23. Transposons
Fall into two general classes with
respect to how they move.
One class encodes proteins that move
the DNA element directly to a new
position or replicate the DNA.
– Found in both prokaryotes and eukaryotes
The other class are related to
retroviruses in that they encode a
reverse transcriptase for making DNA
copies of their RNA transcripts, which
then integrate at new sites in the
genome.
– Found only in eukaryotes. 23
24. Transposable elements are
important because they can
insert into sites where there is
no sequence homology
(nonhomologous
recombination)
24
25. Prokaryotes
What are two types of
transposons in prokaryotes and
how do they differ? (IS and Tn)
– What enzyme is required for the
transposition of an IS element?
– How is a composite transposon
different from a noncomposite
transposon?
– How does the replicative
transposition mechanism differ from
the conservative mechanism of 25
30. EUKARYOTIC TRANSPOSITION
What is cytogenetics, and how was it
used to find “jumping genes” in
eukaryotes?
In what ways are eukaryotic
transposable elements similar to
those found in prokaryotes?
What can determine the stability of a
newly-inserted transposable element
in plants?
30
31. What genes do Ty elements in
yeast carry, and what are their
purposes?
In what ways is the yeast Ty
element similar to a retrovirus?
Why are Ty elements classified
as retroposons?
31
33. Transposable Elements
(Transposons)
DNA elements capable of moving ("transposing")
about the genome
Discovered by Barbara McClintock, largely from
cytogenetic studies in maize, but since found
in most organisms
She was studying "variegation" or sectoring in
leaves and seeds
She liked to call them "controlling elements“
because they affected gene expression in
myriad ways
33
34. 1. Nobelprize.org
(1983 Nobel Prize in
Physiology and Medicine)
2. profiles.nlm.nih.gov/LL/
Barbara McClintock 1902-1992
34
Corn (maize) varieties
35. Corn evolution in 7000 yrs of domestication
cob of Hopi Blue corn
cob of wild teosinte
35
37. Mutant Kernel Phenotypes
1. Pigmentation mutants
– affect anthocyanin pathway
– elements jump in/out of transcription
factor genes (C or R)
– sectoring phenotype - somatic mutations
– whole kernel effected - germ line
mutation
2. Starch synthesis mutants
- stain starch with iodine, see sectoring in
endosperm
37
38. Some maize phenotypes caused by transposable
elements excising in somatic tissues.
Start with lines that produce kernels defective in starch synthesis
(endosperm phenotypes) or anthocyanin synthesis (aleurone and
pericarp phenotypes) because of an inserted element, and the
38
element excises during development.
40. Other Characteristics of McClintock's
Elements
Unstable mutations that revert frequently but often
partially, giving new phenotypes.
Some elements (e.g., Ds) correlated with
chromosome breaks.
Elements often move during meiosis and
mitosis.
Element movement accelerated by genome
damage.
40
41. Molecular Analysis of Transposons
Transposons isolated by first cloning a gene that
they invaded. A number have been cloned this way,
vAia "Transposon trapping“.
Some common molecular features:
– Exist as multiple copies in the genome
– Insertion site of element does not have extensive
homology to the transposon
– Termini are an inverted repeat
– Encode “transposases” that promote movement
– A short, direct repeat of genomic DNA often
flanks the transposon : “Footprint”
41
42. Ac and Ds
Ds is derived from Ac by internal deletions
Ds is not autonomous, requires Ac to move
Element termini are an imperfect IR
Ac encodes a protein that promotes
movement - Transposase
Transposase excises element at IR, and also
cuts the target
42
43. Structure of Ac and Ds deletion
derivatives
Ds is not autonomous, requires Ac to move! 43
Fig. 23.10
44. How duplications
in the target site
probably occur.
Duplication
remains when
element excises,
thus the
Footprint.
44
Fig. 23.2
45. Mu/MuDR (Mutator)
Discovered in maize; differs significantly
from Ac and En/Spm families
Autonomous and non-autonomous
versions; many copies per cell
Contain a long TIR (~200 bp)
Transpose via a gain/loss (somatic
cells) or a replicative (germline cells)
mechanism.
45
46. Structure of MuDR
(autonomous Mu)
and its promoters.
• MuDrA and B
expressed at high
levels in dividing cells
and pollen, because of
transcriptional
enhancers.
• MURA is transposase
& has NLS.
• MURB needed for
insertion in somatic
cells.
46
48. Control of Transposons
Autoregulation: Some transposases
are transcriptional repressors of
their own promoter(s)
e.g., TpnA of the Spm element
Transcriptional silencing: mechanism
not well understood but correlates
with methylation of the promoter
(also methylation of the IRs)
48
49. Biological Significance of Transposons
They provide a means for genomic change
and variation, particularly in response to
stress (McClintock’s "stress" hypothesis)
(1983 Nobel lecture, Science 226:792)
or just "selfish DNA"?
No known examples of an element playing a
normal role in development.
49
50. Transposable elements
AC and DS in maize
– AC encodes transposase,
required to excise DS
50
53. General Features of Transposable Elements
1. Transposable elements are divided into two classes on the basis of their
mechanism for movement:
a. Some encode proteins that move the DNA directly to a new position or replicate the
DNA to produce a new element that integrates elsewhere. This type is found in both
prokaryotes and eukaryotes.
b. Others are related to retroviruses, and encode reverse transcriptase for making DNA
copies of their RNA transcripts, which then integrate at new sites. This type is found
only in eukaryotes.
2. Transposition is nonhomologous recombination, with insertion into DNA that
has no sequence homology with the transposon.
a. In prokaryotes, transposition can be into the cell’s chromosome, a plasmid or a
phage chromosome.
b. In eukaryotes, insertion can be into the same or a different chromosome.
3. Transposable elements can cause genetic changes, and have been involved in
the evolution of both prokaryotic and eukaryotic genomes. Transposons may:
a. Insert into genes.
b. Increase or decrease gene expression by insertion into regulatory sequences.
c. Produce chromosomal mutations through the mechanics of transposition.
Chapter 20 slide 53
54. Transposable Elements in Prokaryotes
1.Prokaryotic examples include:
a. Insertion sequence (IS) elements.
b.Transposons (Tn).
c. Bacteriophage Mu (replicated by
transposition)
Chapter 20 slide 54
55. Insertion Sequences
Animation: Insertion Sequences in Prokaryotes
1. IS elements are the simplest transposable elements found in
prokaryotes, encoding only genes for mobilization and insertion of its
DNA. IS elements are commonly found in bacterial chromosomes and
plasmids.
2. IS elements were first identified in E. coli’s galactose operon,
wheresome mutations’ were shown to result from insertion of a DNA
sequence now called IS1 (Figure 20.1)
3. Prokaryotic IS elements range in size from 768 bp to over 5 kb. Known
E. coli IS elements include:
a. IS1 is 768 bp long, and present in 4–19 copies on the E. coli
chromosome.
b. IS2 has 0–12 copies on the chromosome, and 1 copy on the F plasmid.
c. IS10 is found in R plasmids.
4. The ends of all sequenced IS elements show inverted terminal repeats
(IRs) of 9–41 bp (e.g., IS1 has 2355 of nearly identical sequence).
Chapter 20 slide
bp
57. 5. Integration of IS elements may:
a. Disrupt coding sequences or regulatory regions.
b. Alter expression of nearby genes by the action of IS element
promoters.
c. Cause deletions and inversions in adjacent DNA.
d. Serve as a site for crossing-over between duplicated IS elements.
6. When an IS element transposes:
a. The original copy stays in place, and a new copy inserts randomly into the chromosome.
b. The IS element uses the host cell replication enzymes for precise replication.
c. Transposition requires transposase, an enzyme encoded by the IS element.
d. Transposase recognizes the IR sequences to initiate transposition.
e. IS elements insert into the chromosome without sequence homology (illegitimate
recombination) at target sites (Figure 20.2).
i. A staggered cut is made in the target site, and the IS element inserted.
ii. DNA polymerase and ligase fill the gaps, producing small direct repeats of the
target site flanking the IS element (target site duplications).
f. Mutational analysis shows that IR sequences are the key
Chapter 20 slide 57
59. Transposons
1. Transposons are similar to IS elements, but carry additional genes, and have a
more complex structure. There are two types of prokaryotic transposons:
a. Composite transposons carry genes (e.g., antibiotic resistance) flanked on both sides
by IS elements (IS modules).
i. The IS elements are of the same type, and called ISL (left) and ISR (right).
ii. ISL and ISR may be in direct or inverted orientation to each other.
iii. Tn10 is an example of a composite transposon (Figure 20.3). It is 9.3 kb, and
contains:
(1) 6.5 kb of central DNA with genes that include tetracycline resistance (a
selectable marker).
(2) 1.4 kb IS elements (IS10L and IS10R) at each end, in an inverted
orientation.
iv. Transposition of composite transposons results from the IS elements, which
supply transposase and its recognition signals, the IRs.
(1) Tn10’s transposition is rare, because transpose is produced at a rate of ,1
molecule/generation.
(2) Transposons, like IS elements, produce target site duplications (e.g., a 9-
bp duplication for Tn10). (Table 20.1)
Chapter 20 slide 59
61. b. Noncomposite transposons also carry genes (e.g., drug
resistance) but do not terminate with IS elements.
i. Transposition proteins are encoded in the central
region.
ii. The ends are repeated sequences (but not IS
elements).
iii. Noncomposite transposons cause target site
duplications (like composite transposons).
iv. An example is Tn3.
(1) Tn3’s length is about 5 kb, with 38-bp inverted terminal
repeats.
(2) It has three genes in its central region:
(a) bla encodes β-lactamase, which breaks down ampiciliin.
(b) tnpA encodes transposase, needed for insertion into a new site.
(c) tnpB encodes resolvase, involved in recombinational events needed for
transposition (not found in all transposons).
(3) Tn3 produces 20 slide 61 duplication upon insertion (Figure
Chapter
a 5-bp
20.5).
64. 2. Models have been generated for transposition:
a. Cointegration is an example of the replicative transposition that
occurs with Tn3 and its relatives (Figure 20.6).
i. Donor DNA containing the Tn fuses with recipient DNA.
ii. The Tn is duplicated, with one copy at each donor-recipient DNA
junction, producing a cointegrate.
iii. The cointegrate is resolved into two products, each with one copy
of the Tn.
b. Conservative (nonreplicative) transposition is used by Tn10, for
example. The Tn is lost from its original position when it transposes.
3. Transposons cause the same sorts of mutations caused by IS elements:
a. Insertion into a gene disrupts it.
b. Gene expression is changed by adjacent Tn promoters.
c. Deletions and insertions occur.
d. Crossing-over results from duplicated Tn sequences in the genome.
Chapter 20 slide 64
66. IS Elements and Transposons in Plasmids
1. Bacterial plasmids are extrachromosomal DNA capable of self-replication.
Some are episomes, able to integrate into the bacterial chromosome. The E. coli
F plasmid is an example (Figure 20.7):
a. Important genetic elements of the F plasmid are:
i. tra genes for conjugal transfer of DNA from donor to recipient.
ii. Genes for plasmid replication.
iii. 4 IS elements: 2 copies of IS3, 1 of IS2, and 1 of γδ (gammadelta). All have
homology with IS elements itt the E. coli chromosome.
b. The F factor integrates by homologous recombination between IS elements,
mediated by the tra genes.
2. R plasmids have medical significance, because they carry genes for resistance to
antibiotics, and transfer them between bacteria (Figure 20.7).
a. Genetic features of R plasmids include:
i. The resistance transfer factor region (RTF), needed for conjugal transfer. It
includes a DNA region homologous to an F plasmid region, and genes for
plasmid-specific DNA replication.
ii. Differing sets of genes, such as those for resistance to antibiotics or heavy
metals. The resistance genes are transposons, flanked by IS module-like
sequences, and can replicate and insert into the bacterial chromosome.
b. R plasmids are clinically significant, because they disseminate drug resistance genes
between bacteria.
Chapter 20 slide 66
68. Bacteriophage Mu
1. Temperate bacteriophage Mu (mutator) can cause mutations when it transposes.
Its structure includes:
a. A 37 kb linear DNA in the phage particle that has central phage DNA and unequal
lengths of host DNA at the ends (Figure 20.8).
b. The DNA’s G segment can invert, and is found in both orientations in viral DNA.
2. Following infection, Mu integrates into the host chromosome by conservative
(non-replicative) transposition.
a. Integration produces prophage DNA flanked by 5 bp target site direct repeats.
b. Flanking DNA from the previous host is lost during integration.
c. The Mu prophage now replicates only when the E. coli chromosome replicates, due
to a phage-encocled repressor that prevents most Mu gene expression.
3. Mu prophage stays integrated during the lytic cycle, and replication of Mu’s
genome is by replicative transposition.
4. Mu causes insertions, deletions, inversions and translocations (Figure 20.9).
Chapter 20 slide 68
71. Transposable Elements in Eukaryotes
1. Rhoades (1930s) working with sweet corn, observed interactions between two
genes:
a. A gene for purple seed color, the Al locus. Homozygous mutants (a/a) have colorless
seeds.
b. A gene on a different chromosome, Dt (dotted) that causes seeds with genotype a/a
Dt/-- to have purple dots.
i. Dt appears to mutate the a allele back to the Al wild-type in regions of the seed,
producing a dotted phenotype.
ii. The effect of the Dt allele is dose dependent.
(1) One dose gave an average of 7.2 dots per seed.
(2) Two doses gave an average of 22.2 dots/seed.
(3) Three doses gave an average of 121.9 dots/seed.
c. Rhoades interpreted Dt as a mutator gene.
2. McClintock (1940s-50s), working with corn (Zea mays) proposed the existence
of “controlling elements” that regulate other genes and are mobile in the
genome.
3. The genes studied by both Rhoades and McClintock have turned out to be
transposable elements, and many others have been identified in various
eukaryotes.
a. Most studied are transposons of yeast, Drosophila, corn and humans.
b. Their structure is very similar to that of prokaryotic transposable elements.
c. Eukaryotic transposable elements have genes for transposition and integration at a
number of sites, as well as a variety of other genes.
d. Random insertion results from 20 slide 71
Chapter non-homologous recombination, and means that any
chromosomal gene may be regulated by a transposon.
72. Transposons in Plants
Animation: Transposable Elements in Plants
1. Plant transposons also have IR sequences, and generate short direct target site repeats.
2. The result of transposon insertion into a plant chromosome will depend on the properties
of the transposon, with possible effects including:
a. Activation or repression of adjacent genes by disrupting a cellular promoter, or by action of
transposon promoters.
b. Chromosome mutations such as duplications, deletions, inversions, translocations or breakage.
c. Disruption of genes to produce a null mutation (gene is nonfunctional).
3. Several families of transposons have been identified in corn, each with characteristic
numbers, types and locations.
a. Each family has two forms of transposon. Either can insert into a gene and produce a mutant
allele.
i. Autonomous elements, which can transpose by themselves. Alleles produced by an
autonomous element are mutable alleles, creating mutations that revert when the
transposon is excised from the gene.
ii. Nonautonomous elements, which lack a transposition gene and rely on the presence of
another transposon to supply the missing function. Mutation by these elements is stable
(except when an autonomous element from the family is also present).
Chapter 20 slide 72
73. 4. Multiple genes control corn color, and classical genetics indicates that a
mutation in any of these genes leads to a colorless kernel. McClintock
studied the unstable mutation that produces spots of purple pigment on
white kernels (Figure 20.10).
a. She concluded that spots do not result from a conventional mutation,
but from a controlling element (now Tn).
b. A corn plant with genotype c/c will have white kernels, while C/-- will
result in purple ones.
i. If a reversion of c to C occurs in a cell, that cell will produce purple
pigment, and hence a spot.
ii. The earlier in development the reversion occurs, the larger the spot.
Chapter 20 slide 73
74. iii. McClintock concluded that the c allele resulted from
insertion of a “mobile controlling element” into the C allele.
(1) The element is Ds (dissociation), now known to be a
nonautonomous transposon.
(2) Its transposition is controlled by Ac (activator), an
autonomous transposon (Figure 20.11).
c. McClintock’s evidence of transposable elements did not fit the
prevailing model of a static genome. More recent studies have
confirmed and characterized the elements involved.
i. The Ac-Ds system involves an autonomous element (Ac)
whose insertions are unstable, and a nonautonomous element
(Ds) whose insertions are stable if only Ds is present.
ii. McClintock (1950s) showed that some Ds elements derive
from Ac elements.
Chapter 20 slide 74
76. iii. Ac is 4,563 bp, with 1 1-bp imperfect terminal IRs and 1 transcription
unit producing a 3.5 kb mRNA encoding an 807 amino acid transposase.
Insertion generates an 8-bp target site duplication (Figure 20.12).
iv. Ac activates Ds to transpose or break the chromosome where it is
inserted.
v. Ds elements vary in length and sequence, but all have the same terminal
IRs as Ac, and many are deleted or rearranged versions of Ac.
vi. Unique to corn transposons, timing and frequency of transposition and
gene rearrangements are developmentally regulated.
vii. Ac transposes only during chromosome replication, and does not leave a
copy behind. There are two possible results of Ac transposition,
depending on whether the target DNA has replicated or not (Figure
20.13). -
(1) If Ac transposes during replication into a replicated target site, its
chromatid’s donor site will be empty since that copy of Ac has
inserted elsewhere. In the homologous donor site on the other
chromatid, a copy will remain. There is no net increase in copies of
Ac.
(2) Transposition to an unreplicated chromosome site also leaves one
donor site empty (and the other with a copy of Ac). The DNA into
which Ac inserts will then be replicated, resulting in a net gain of one
copy of Ac.
viii. Replication of Ds is the same, except that the transposition protein is
supplied by an integrated 20 slide 76
Chapter Ac element.
79. 5. In Mendel’s wild-type (SS) peas the starch grains are
large and simple, while in wrinlded peas (ss) they are
small and fissured.
a. SS seeds contain more starch and less sucrose than ss seeds.
b. The sucrose difference makes ss seeds larger, with higher water
content, so that when dried they are wrinided.
c. One type of starch-branching enzyme (SBEI) is missing in ss
plants, reducing their starch content.
d. The SBEI gene corresponding to the s allele has a 0.8 kb
transposon similar to the Ax/Ds family inserted into the wild-
type S allele.
Chapter 20 slide 79
80. Ty Elements in Yeast
1. Ty elements share characteristics with bacterial transposons:
a. Terminal repeated sequences.
b. Integration at non-homologous sites.
c. Generation of a target site duplication (5 bp).
2. Ty element is diagrammed in Figure 20.14:
a. It is 5.9 kb including 2 terminal direct repeats of 334 bp, the long terminal repeats
(LTR) or deltas (δ).
b. Each delta contains a promoter and transposase recognition sequences.
c. Ty elements encode one 5.7 kb mRNA beginning at the delta 5’ promoter (Figure
20.14).
d. There are two ORFs in the mRNA, designated TyA and TyB, encoding two different
proteins.
e. Ty copy number varies between yeast strains, with an average of about 35.
3. Ty elements also share similarities with retroviruses, ssRNA viruses that replicate
via dsDNA intermediates.
a. Ty elements transpose by making an RNA copy of the integrated DNA sequence, them
making DNA using reverse transcriptase. This DNA can integrate at a new
chromosomal site. Evidence for this includes:
i. An experimentally introduced intron in the Ty element (which normally lacks
introns) was monitored through transposition. The intron was removed, indicating
an RNA intermediate.
ii. Ty elements encode a reverse transcriptase.
iii. Virus-like particles containing Ty RNA and reverse transcriptase activity occur.
Chapter 20 slide 80
b. Ty elements are referred to as retrotransposons.
82. Drosophila transposons
1. It is estimated that 15% of the Drosophila genome is mobile! These
transposons fall into different classes:
a. The copia retrotransposons include several families, each highly
conserved and present in 5-100 widely scattered copies per genome
(Figure 20.15).
i. All copia elements in Drosophila can transpose, and there are
differences in number and distribution between fly strains.
ii. Structurally, copia elements are similar to yeast Ty elements:
(1) Direct LTRs of 276 bp flank a 5 kb DNA segment.
(2) The end of each LTR has 17 bp inverted repeats.
(3) An RNA intermediate and reverse transcriptase are used for
transposition.
(4) Virus-like particles (VLPs) occur with copia.
(5) Integration results in target site duplication (3-6 bp).
Chapter 20 slide 82
84. b. P elements cause hybrid dysgenesis, a series of
defects (mutations, chromosomal aberrations and
sterility) that result from crossing certain
Drosophila strains (Figure 20.16).
i. A mutant lab strain female (M) crossed with a wild-type
male (P) will result in hybrid dysgenesis.
ii. A mutant lab strain male (M) crossed with a wild-type
(P) female (reciprocal cross) will have normal offspring.
iii. Thus, hybrid dysgenesis results when chromosomes of
the P male parent enter cytoplasm of an M type oocyte,
but cytoplasm from P oocytes does not induce hybrid
dysgenesis.
Chapter 20 slide 84
86. iv. The model is based on the observation that the M strain has no P
elements, while the haploid genome of the P male has about 40
copies.
(1) P elements vary from full-length autonomous elements
through shorter versions resulting from a variety of internal
deletions.
(2) P element transposition is activated only in the germ line.
(3) The F1 of an M female crossed with a P male have P
elements inserted at new sites, flanked by target site repeats.
(4) P elements are thought to encode a repressor protein that
prevents transposase gene expression, preventing transposition.
(5) Cytoplasm in an M oocyte lacks the repressor, and so when
fertilized with P-bearing chromosomes, transposition occurs
into the maternal chromosomes, leading to hybrid dysgenesis.
v. P elements are used experimentally to transfer genes into the germ
line of Drosophila embryos. For example (Figure 20.18):
(1) The wild-type rosy (ry) gene was inserted into a P element,
cloned in a plasmid and microinjected into a mutant ry/ry strain.
(2) Insertion of the recombinant P element into the recipient
chromosome introduced the ry allele, and produced wild-type
flies.
Chapter 20 slide 86
89. Human Retrotansposons
1. Retrotransposons also appear to be present in mammals. For example, a very
abundant human SINE repeat (short interspersed sequence) is the Mu
family, named for the AluI restriction site in its sequence.
a. Mu sequences are about 300 bp, repeated 300,000-500,000 times in the human
genome (up to 3% of total human DNA).
b. Sequences are divergent, related but not identical.
c. Each Mu sequence is flanked by 7-20 bp direct repeats.
d. At least a few Mu sequences can be transcribed, and the model is that
transcriptionally active Mu sequences are retrotransposons that move via an
RNA intermediate.
e. A human case of a genetic disease, neurofibromatosis, provides some evidence.
i. Neurofibromas (tumorlike growths on the body) result from an autosomal
dominant mutation.
ii. In a patient’s DNA, an unusual Mu sequence was detected in one of the
introns of the neurofibromatosis gene.
iii. The resulting longer transcript is incorrectly proessed, removing an exon
from the mRNA and producing a nonfunctional protein.
iv. Neither parent had this Mu sequence in the neurofibromatosis gene.
v. Divergent Mu sequences made it possible to track this particular version to
an insertion event in the germ line of the patient’s father.
f. It is not clear how the functions needed for Mu retrotransposition are provided.
Chapter 20 slide 89
90. 2. A mammalian LINEs family, LINEs-i (Li elements) is
also thought to be retrotransposons.
a. Humans have 50,000-100,000 copies of the Li element,
comprising about 5% of the genome.
b. The full-length element (6.5 kb) is not abundant, and most Li
elements are deleted versions.
c. The full-length Li element contains a large ORF with homolegy
to known reverse transcriptases. Experimentally, the Li ORF can
substitute for the yeast Ty reverse transcriptase gene.
d. Li elements are thought to be retrotransposons, but do not have
LTRs.
e. Clinically, cases of hemophilia have been shown to result from
newly transposed Li insertions into the factor VIII gene. (Factor
VIII is required for normal blood clotting.)
Chapter 20 slide 90
92. 21.2 Insertion Sequences
Are Simple Transposition
Modules
An insertion sequence is a
transposon that codes for the
enzyme(s) needed for
transposition flanked by short
inverted terminal repeats.
92
93. The target site at
which a transposon
is inserted is
duplicated during
the insertion
process.
– This forms two
repeats in direct
orientation at the
ends of the
transposon.
The length of the
direct repeat is:
– 5 to 9 bp
Figure 21.2 93
94. 21.3 Composite
Transposons Have IS
Modules
Transposons can carry other
genes in addition to those coding
for transposition.
Composite transposons have a
central region flanked by an IS
element at each end.
94
95. Either one or both of
the IS elements of a
composite
transposon may be
able to undertake
transposition.
A composite
transposon may
transpose as a unit.
– An active IS element
at either end may also
transpose
Figure 21.3
95
96. by Both Replicative and
Nonreplicative
Mechanisms
All transposons
use a common
mechanism in
which:
– staggered nicks
are made in target
DNA
– the transposon is
joined to the
protruding 96
ends
– the gaps are filled
Figure 21.5
97. The order of events and exact
nature of the connections between
transposon and target DNA
determine whether transposition
is:
– replicative
– nonreplicative
Figure 21.6 Figure 21.7 97
98. 21.5 Transposons Cause
Rearrangement of DNA
Homologous recombination between
multiple copies of a transposon
causes rearrangement of host DNA.
Homologous recombination between
the repeats of a transposon may lead
to precise or imprecise excision.
98
99. 21.6 Common
Intermediates for
Transposition
Transposition starts by
forming a strand
transfer complex.
– The transposon is
connected to the target
site through one strand
at each end.
99
Figure 21.11
100. The Mu transposase
forms the complex by:
– synapsing the ends of Mu
DNA
– followed by nicking
– then a strand transfer
reaction
Replicative
transposition follows if
the complex is
replicated. 100
Figure 21.12
101. 21.7 Replicative
Transposition Proceeds
through a Cointegrate
Replication of a strand
transfer complex
generates a
cointegrate:
– A fusion of the donor
and target replicons.
The cointegrate has
two copies of the
transposon.
– They lie between the
101
Figure 21.13
102. Recombination between the
transposon copies regenerates the
original replicons, but the recipient
has gained a copy of the transposon.
The recombination reaction is
catalyzed by a resolvase coded by the
transposon.
102
103. 21.8 Nonreplicative
Transposition Proceeds
by Breakage and Reunion
Nonreplicative transposition results if:
– a crossover structure is nicked on the
unbroken pair of donor strands and
– the target strands on either side of the
transposon are ligated
103
Figure 21.15
104. Two pathways for nonreplicative
transposition differ according to
whether:
– the first pair of transposon strands are
joined to the target before the second
pair are cut (Tn5), or
– whether all four strands are cut before
joining to the target (Tn10)
104
105. 21.9 TnA Transposition
Requires Transposase
and Resolvase
Replicative transposition of TnA requires:
– a transposase to form the cointegrate structure
– a resolvase to release the two replicons
The action of the resolvase resembles
lambda Int protein.
It belongs to the general family of
topoisomerase-like site-specific
recombination reactions.
– They pass through an intermediate in which the
105
106. 21.10 Transposition of
Tn10 Has Multiple
Controls
Multicopy inhibition reduces the rate of
transposition of any one copy of a transposon
when other copies of the same transposon are
introduced into the genome.
Multiple mechanisms affect the rate of
transposition.
106
Figure 21.21
107. Elements in Maize Cause
Breakage and
Rearrangements
Transposition in maize was
discovered because of the effects of
chromosome breaks.
– The breaks were generated by
transposition of “controlling elements.”
The break generates one
chromosome that has:
– a centromere
– a broken end 107
– one acentric fragment
108. The acentric
fragment is lost
during mitosis;
– this can be
detected by the
disappearance of
dominant alleles
in a heterozygote.
Figure 21.23
108
109. Fusion between the
broken ends of the
chromosome generates
dicentric chromosomes.
– These undergo further
cycles of breakage and
fusion.
The fusion-breakage-
bridge cycle is
responsible for the
occurrence of somatic
variegation.
109
Figure 21.24
110. 21.12 Controlling
Elements Form Families
of Transposons
Each family of
transposons in
maize has both
autonomous and
nonautonomous
controlling
elements.
110
Figure 21.25
111. Autonomous controlling elements
code for proteins that enable them to
transpose.
Nonautonomous controlling elements
have mutations that eliminate their
capacity to catalyze transposition.
– They can transpose when an autonomous
element provides the necessary proteins.
Autonomous controlling elements
have changes of phase, when their
properties alter as a result of changes
111
112. 21.13 Spm Elements
Influence Gene
Expression
Spm elements affect gene
expression at their sites of
insertion, when the TnpA protein
binds to its target sites at the
ends of the transposon.
Spm elements are inactivated by
methylation.
112
113. 21.14 The Role of
Transposable Elements in
Hybrid Dysgenesis
P elements are transposons that are
carried in P strains of Drosophila
melanogaster, but not in M strains.
When a P male is crossed with an M
female, transposition is activated.
113
114. The insertion of P elements at new sites in
these crosses:
– inactivates many genes
– makes the cross infertile
114
Figure 21.28
115. 21.15 P Elements Are
Activated in the Germline
P elements are
activated in the
germline of P male x
M female crosses.
This is because a
tissue-specific
splicing event
removes one intron.
– This generates the
coding sequence for
Figure 21.29
115
116. The P element also
produces a
repressor of
transposition.
– It is inherited
maternally in the
cytoplasm.
The presence of the
repressor explains
why M male x P
Figure 21.30 female crosses
remain fertile.
116
117. Pray, L. (2008) Transposons: The
jumping genes. Nature
Education 1(1)
118. DNA transposons
Seen in both prokaryotes and
eukaryotes
– the IS element (insertion sequence) in bacteria
– DNA transposons seen in eukaryotic genomes
(P elements in fruit flies, Ac/Ds elements in
plant genomes)
Mechanism of transposon
action
– Transposon encodes an enzyme: transposase
– Transposase excises itself out and then is able
to cut in the middle of a target DNA
– Effect is based on where the transposable
119. RNA transposable
elements
Derived from an RNA
intermediate
Seen only in eukaryotic genomes
Originated from ancient retroviral
genome
– Retrotransposon
LTR elements
– Retroposons
SINE-human
LINE-human
120. - Derived from a viral genome from the
retrovirus:
LTR gag RT env LTR
RT: reverse transcriptase ~7 kb
LTR: long terminal repeat
gag, env: encode proteins needed for retroviral assembly and
infection
Retroelements: missing some or most of the complete retroviral
genome;
122. Mechanism of retrotransposition
RNA
Retrotransposon
Transcription
Reverse transcription
RNA
DNA
Retrotransposon
Retrotransposon copy
123. Human Retroposons: non-LTR
- LINE: long interspersed elements
gag? RT poly(A)
~6 kb
-SINE: short interspersed element;
The Alu element is a well known example
poly(A)
~0.3 kb
124. C-value paradox: genome size not
always predictor of gene number
Taken fron http://cs.uni.edu
126. Types of Transposable
Elements
Type Transposition Examples
Transposon Use a DNA Corn: Ds element
(Class I) intermediate Drosophila: P
element
Retrotransposons Use an RNA Yeast: Ty
(Class II) intermediate Drosophila: Copia
Human: Alu
Human: L1
Transposition: movement of a transposable element
127. Characteristics of Transposable
Elements
All elements
have
direct repeats:
short repeated
sequences
flanking the
element, arise
by transposition
129. Characteristics of Transposable
Elements
Carry gene for enzyme that
catalyzes transposition
– transposase for elements that
use a DNA intermediate
– reverse transcriptase for
elements that use an RNA
intermediate
May contain other genes
130. Mechanisms of Transposition
Use of a DNA Intermediate
– Replicative- new copy in new
location, old copy retained at
original site, element is used as
template to produce the new copy
131. Mechanisms of Transposition
Use of a DNA Intermediate
– Non-replicative: moves to another
site without replication of the
element
132. Mechanisms of Transposition
Use of an RNA
Intermediate
– element is
transcribed
– reverse
transcriptase
produces a
double-stranded
DNA copy for
insertion at
another site
133. Types of Retrotransposons
Viral Retrotransposons
– resemble retroviruses = viruses with
an RNA genome
Long terminal direct repeat at each end
Carry genes for enzymes usually found in
RNA viruses
135. Types of Retrotransposons
Non-viral Retrotransposons
– do not resemble retroviruses
– two types in humans
LINES = long interspersed elements
– 6-7 kb long
– example: L1 has 600,000 copies, represents
15% of human DNA
SINES = short interspersed elements
– 300 bp long
– example: Alu has 1 million copies, represents
10% of human DNA
136. Definitions and Keywords
Transposons - are sequences of DNA that can move
around to different positions within the genome of a
single cell, a process called transposition.
Transposase -An enzyme that binds to ends of
transposon and catalyses the movement of the
transposon to another part of the genome by a cut and
paste mechanism or a replicative transposition
mechanism.
IS elements -A short DNA sequence that acts as a
simple transposable element
137. Definitions and
Keywords
DNA polymerase-A DNA
polymerase is an enzyme that
catalyzes the polymerization of
deoxyribonucleotides into a
DNA strand.
DNA ligase is a special type of
ligase that can link together
two DNA strands that have
double-strand break a break in
138. Bacterial Transposons
Bacteria contain two types of transposons
1]Composite mobile genetic elements that are larger than IS
elements and contain one or more protein-coding genes in
addition to those required for transposition.
2]Non composite mobile genetic elements are those which
lack IS elements on its ends e.g. is Tn3
141. Transposable Elements
They are discrete sequence in the
genome that are mobile
they are able to transport themselves
to other location. Other names:
Jumping genes
Selfish DNAs
Molecular parasites
Controlling elements
TEs are present in the genome all species of three domains
142. What do we want to know
about mobile genetics
elements?
1 – The history of mobile genetic
elements
2 – The classification of TEs
3 – The structure of TEs
4 – The mechanism of transposition
5 – The effects of TEs on gene and
genome
6 – The use of TEs as molecular
144.
TRANSPOSONS
“Transposable elements”
“Jumping genes”
Mobile DNA
– able to move from one place to
another within a cell’s genome
– sometimes a copy is made and the
copy moves
– insertion requires target DNA
sequences
146. In the process, they may
- cause mutations.
- increase (or decrease) the
amount of DNA in the
genome.
- promote genome
rearrangements.
- regulate gene expression.
- induce chromosome
breakage and
147. Discovery of
transposons
Barbara McClintock 1950’s Ac Ds
system in maize influencing kernel
color unstable elements
changing map position promote
chromosomal breaks.
Rediscovery of bacterial insertion
sequences
source of polar mutations discrete
change in physical length of DNA
inverted repeat ends: form
“lollipops” in EM after denaturation.
148. These mobile segments of DNA are
sometimes called "jumping genes"
There are two distinct types of
transposons:
1) DNA transposons
-transposons consisting only of DNA
that moves directly from place to place
2) Retrotransposons
- first transcribe the DNA into RNA and
then
- use reverse transcriptase to make a
DNA copy of the RNA to insert in a
new location
149. Classification of Transposons into two classes
In both cases
ds
DNA
intermediate
is integrated
into
the target site in
DNA to complete
150. BACTERIAL TRANSPOSONS
In bacteria, transposons can
jump from chromosomal DNA to
plasmid DNA and back.
Transposons in bacteria usually
carry an additional gene for
function other than transposition---
often for antibiotic resistance.
Bacterial transposons of this
type belong to the Tn family. When
the transposable elements lack
additional genes, they are known
152. 1.Insertion sequences
Insertion sequences – IS1 and IS186,
present in the 50-kb segment of the E.
coli DNA, are examples of DNA
transposons.
Single E. coli genome may contain 20
of them.
Most of the sequence is taken by one or
two genes for transposase enzyme that
catalyses transposition.
IS elements transpose either replicatively
153. cont….
IS elements
Study of E. coli mutations resulting from insertion of 1-
2 kb long
sequence in the middle of certain genes.
Inserted stretches or insertion sequences – could be
visualized by EM.
IS - molecular parasites in bacterial cells.
Transposition of IS is very rare – one in 105-107 cells
per generation.
Higher rates result in greater mutation rates.
154. Bacterial IS element
Central region encodes for one or two enzymes required for transposition. It
is flanked by inverted repeats of characteristic sequence.
The 5’ and 3’ short direct repeats are generated from the target-site DNA
during the insertion of mobile element.
The length of these repeats is constant for a given IS element, but their
sequence depends upon the site of insertion and is not characteristic for the
IS element.
Arrows indicate orientation.
155. Insertion sequences in E.coli
Elements Size (bp) No.of.copies/
genome
IS1 768 8
IS2 1327 5
IS3 1300 1 or more
IS4 1426 1 or more
156.
157. 2.Composite transposons
Bacteria contain composite mobile
genetic elements that are larger than IS
elements and contain one or more
protein-coding genes in addition to those
required for transposition:
Composite transposons - are basically
the pair of IS elements flanking a
segment of DNA usually containing one or
more genes, often coding for AB
resistance.
158. Cont…
2.Composite transposon
- Antibiotic resistant gene
- Flank by IS element
(inverted or directed repeat)
- Terminal IS can transpose by in self
Ex. Tn5, Tn9, Tn10
159.
160. 3. Tn 3 transposon
family
- 5000 bp
- code for Transposase, β-
lactamase,
Resolvase
- Function of resolvase
Decrease Transposase
production
Catalyse the
recombination of
163. 4.Transposable phage
Transposable phages –
bacterial viruses which
tranpose replicatively as a part
of their normal infectious
cycle.
Integrate into E. coli
chromosome at regulatory
element
166. Mechanism of
transposition
Two distinct mechanisms of
transposition:
Replicative transposition – direct
interaction between the donor
transposon and the target site,
resulting in copying of the donor
element
Conservative transposition –
167. Mechanism of transposition
1. Replicative transposition
Copy of transposon sequence
Transposase enzyme cut target DNA
Transposition
Duplication of target sequence
169. 2. Non-replicative
(conservative)
transposition
- Cannot copy transposon
sequence
- Transposition by cut and paste
model
Cut transposon sequence
from donor molecule
attach to target site
172. Evolution of
Transposons
Transposons are found in all
major branches of life.
It arisen once and then spread
to other kingdoms by
horizontal gene transfer.
Duplications and DNA
rearrangements contributed
greatly to the evolution of new
173. Cont…
Mobile DNA most likely also
influenced the evolution of genes
that contain multiple copies of
similar exons encoding similar
protein domains (e.g., the
fibronectin gene).
The evolution of an enormous
variety of antibiotic resistance
transposons and their spread
among bacterial species.
example of genetic adaptation
174. Transposons causing
diseases
Transposons are mutagens. They can damage the
genome of their host cell in different ways:
1. A transposon or a retroposon that inserts
itself into a functional gene will most likely
disable that gene.
2.After a transposon leaves a gene, the resulting
gap will probably not be repaired correctly.
3.Multiple copies of the same sequence, such as
Alu sequences can hinder precise chromosomal
pairing during mitosis and meiosis, resulting in
unequal crossovers, one of the main reasons for
chromosome duplication.
175. Cont…
Diseases caused by
transposons include
-hemophilia A and B
-severe combined
immunodeficiency
-Porphyria
-Cancer
176. Applications
The first transposon was discovered in
the plant maize (Zea mays, corn species),
and is named dissociator (Ds).
Likewise, the first transposon to be
molecularly isolated was from a plant
(Snapdragon).
Transposons have been an especially
useful tool in plant molecular biology.
Researchers use transposons as a means
of mutagenesis.
177. Cont…
To identifying the mutant allele.
To study the chemical
mutagenesis methods.
To study gene expression.
Transposons are also a widely used
tool for mutagenesis of most
experimentally tractable
179. Why study mobile genetic
elements?
They are the major forces
driving evolution
They can cause genome
rearrangement (mutation ,
deletion and insertion )
They have wide range of
application potentials
180. The discovery of mobile
genetic elements
Transposable elements
Phage
Plasmid DNA
181. The discovery of transposable elements
Barbara Mc Clintock discovered TEs in
maize (1983)
Her work on chromosome breakage
began by investigating genetic
instability (1983)
Observing variegated patterns of
pigmentation in maize plant and kernels
New kinds of genetic instability
She spent the next tree decades for this
genetic elements
Controlling elements (1956)
183. Plasmid , phage
Cell to cell conjugation
Bactriophage mediated
transduction
Bill Hayes ( 1952 )
Ellin Wollman and Francois
Jancob , 1961
Alan Campbell
184. Classification of
transposable elements
DNA transposons
Retrotransposons
185. Autonomous and non autonomous
elements
Both class are subdivided into distinct
superfamilies and families
Structure feature , internal organization , the
size of target site duplication , sequence
similarities at the DNA and protein levels
Autonomous : they have the ability to excise
and transpose
non autonomous elements
- They don’t transpose
- They become unstable only when an
autonomous member of same family is
present elsewhere in the genome
- They are derived from autonomous elements
A family consists of single type of autonomous
element accompanied by many varieties of
non autonomous elements
186.
187. DNA based elements
Insertion sequence (IS)
The simplest (smallest)
transposons are called IS
The IS elements are normal
constituents of bacterial
chromosome and plasmids
Spontaneous mutation of the lac
and gal operons
They are autonomous units ,each
of which codes only transposase
189. Composite
transposone
One class of large
transposons are called
Composite transposons
They carring the druge
marker is flanked on
either side by arms that
consist of IS elements
IS modules- identical (both
functional: Tn9; Tn903) or
closely related (differ in
functional ability: Tn10; Tn5)
1. A functional IS module
can transpose either itself or
the entire transposon
190. Mechanism of
transposition
Direct repeats are
generated by
introduction of
staggered cuts whose
protruding ends are
linked to the transposon.
The stugger
between the cuts
determines the
length of the
direct repeats.
The target
repeat is
characteristic
of each
transposon;
reflects the
191. Mechanism of transposition
1- Replicative transpositon
1. Replicative :
a) Transposon is duplicated; a copy of the original element is made
at a recipient site(TnA); donor keeps original copy
b) Transposition- an increase in the number of Tn copies
c) ENZs: transposase (acts on the ends of original Tn) and resolvase
(acts on the duplicated copies)
192. Mechanism of transposition
2 -Nonreplicative
Nonreplicative :
Transposon moves from one site to another and is
conserved; breaks in donor repaired
b) IS and Tn10 and Tn5 use this mechanism; no Tn
copy increase
c) ENZs: only transposase
193. Donor cut
The first stages of Both
replicative and non-replicative
transpositio are semilar
IS elements, prokaryotic
eukaryotic transposons, and
1. Synapsis stage- two .ends of
bacteriophage Mu
transposon are brought together
2. Transposon nicked at both ends; target
nicked at both strands
3.. Nicked ends joine crosswise;covalent
connection between the transposon the
target
194. Mu integrates by nonreplicative transposition; during lytic
cycle- number of copies amplified by replicative
transposition
22 bp
- MuA binds to ends as
tetramer forming a synapsis.
- MuA subunits act in trans to cut
next to R1 and L1 (coordinately;
two active sites to manipulate cuts in transfers
trans in trans
DNA).
- MuA acts in trans to cut the
target site DNA and mediate
in trans strand transfer
195. The chemistry of Donor and
target cut
OH
OH
OH
Cutting of end only
OH
Cutting of both ends
The 3’-ends ends groups released from flanking
DNA by donor cut reaction
They are nuclophile that attack phosphodiester
bonds in target DNA
196. The product of these
reaction is strand
transfer complex transposon target
In strand transfer
complex transposon is
connected to the target
site through one strand
at each end
Next step differs and
determines the type of
transposition:
Strand transfer complex
can be target for
replication (replicative
transposition) or for
repair (nonreplicative
transposition; breakage
& reunion)
Strand transfer complex
197. Molecular mechanism of transposition (I)
Replicative
transposition
Replicative
transposition
proceeds through a
cointegrate.
Transposition may fuse a donor and
recipient replicon into a cointegrate.
Resolution releases two replicons-each
has copy of the transposon
198. transposition
Donor and target cut
Ligation to
target ends
3. 3’-ends prime replication
The crossover structure contains a single stranded
region at each of the staggered ends=
pseudoreplication forks that provide template for
DNA synthesis cointegrate.
200. Retrotransposon (
retroposons )
Use of an RNA
Intermediate
– element is
transcribed
– reverse
transcriptase
produces a
double-stranded
DNA copy for
insertion at
another site
– they as other
elements
202. retrovirus
RNA
reverstranscriptase
Liner DNA
Integration
provirus
Transcription
RNA
203. LTR - retrotrasposon
pol
Reverse transcriptase (RT)
Integrase (IN)
Ribonuclease H (RH)
gag
?
env
204. mechanism of transposition
Integrase acts on both the retrotransposon line DNA and target D
The integrase bring the ends
of the linear DNA together 5’-ends
-Generate 2 base recessed 3’ -ends 3’-ends
and staggered end in target DNA
205. Non – LTR retrovirus
LINES = long interspersed elements
SINES = short interspersed elements
don’t terminate in the LTRs
they are significant part of relatively
short sequence of mammalian
genomes .
206. Effect of transposabli elements on gene and
genome
TEs cause a varity of change in the
genome of their hosts
this ability to induce mutation depend
on their of capability of transposing
some arrangement can be beneficial
they can advantageous for adaptation
to new environment
play important role in evolution .
they have the ability to rearrange
genomic information in several ways
1 – Modification of gene expression
2 – Alternation gene sequence
3 – Chromosomal structural changes
207. Modification of gene
expression
insertion of a TE within or
adjacent to a gene
the element blocks or alters
the pattern of transcription .
insertion of Fot1 in a intron of niad
(F . oxysporum )
different mutant transcripts all
were shorter
They result from:
208. Alternation gene sequence
cut and pate mechanism often
produce variation when they excise
.
the excision process may result in
addition of a few base pair (
footprint ) at donor site .
these footprint cause
diversification of nucleotide
sequence and new functional
alleles
Example :Fot1 and Impala generally leave 4 bp – (
108 ) or 5 – ( 63 ) foot prints
209. Chromosomal
structural changes
TEs can produce a series of
genome rearrangment through
ectopic recombination
deletion , duplication , inversion
and translucation mediate by TEs (
Drosophila , Yeast , human )
karyoptypic variation in natural
isolate in fungai
high level of chromosome – length
polymorphism (Magnoporthe grisea
, F. oxysporum)
translocation tox1 of Cochliobolus
210. Use of TEs as molecular tools
Use as strain specific
diagnostic tools
TEs are often restricted to
specific strains
identify specific pathogen in
plant pathology
Fot1 ( F. oxysporum f sp.
albedians ) provide PCR targets
a sensitive detection thechnique
to prevent the introduction of
pathogenic form
211. Use of TEs as molecular
tools
Tools for the analysis of population structure
MGR 586 ( Magneporthe
grisea )
oryza : 30 – 50 wheat and
other ( 1 – 2 )
they have used to distinguish
genetically divergent
population
fingerprinting of isolates
212. Gene taggingas molecular tools
Use of TEs with transposable
elements
jumping into coding region
Disrupt target
gene
Target gene can easily
determined by PCR arise mutant
methods phenotype
214. A composite transposon, is flanked by two
Composite Transposon
separate IS elements which may or may not
be exact replicas. Instead of each IS element
moving separately, the entire length of DNA
spanning from one IS element to the other is
transposed as one complete unit.
IR IR
215. Non composite Transposon
Non-composite transposons (which lack flanking
insertion sequences). In each case, transposition
requires specific DNA sequences located at the ends
(IS1, IS3, Tn5, Tn10, and Tn3) or a multisubunit
complex (e.g. Tn7).
Encode transposition proteins, have inverted repeats
(but no ISs) at their ends. In addition to resistance and
virulence genes they may encode catabolic enzymes
216. Mechanism of transposition
There are two mechanisms of transposition replicative
and nonreplicative
During transposition, the IS-element transposase
makes cuts at the positions indicated by small red
arrows,
So the entire transposon is moved from the donor DNA
(e.g., a plasmid).
A DNA polymerase fills in the resulting gaps from the
sticky ends and DNA ligase closes the sugar-phosphate
backbone. This results in target site duplication and
the insertion sites of DNA transposons may be
identified by short direct repeats (a staggered cut in
the target DNA filled by DNA polymerase) followed by
inverted repeats (which are important for the
transposon excision by transposase). The duplications
at the target site can result in gene duplication and this
is supposed to play an important role in evolution.
Composite transposons will also often carry one or
217. Mechanism of
transposition(contd)
The conservative mechanism, also called the “cut-and-paste”
mechanism, is used by elements like Tn10 .
The element is excised cleanly by double-strand cleavages from the
donor DNA
and inserted, with limited repair, between a pair of staggered nicks at
the target DNA.
Replicative transposition is a mechanism of transposition in molecular
biology in which the transposable element is duplicated during the
reaction, so that the transposing entity is a copy of the original element.
Replicative transposition is characteristic to retrotransposons and
occurs from time to time in class II transposons.
Retrieved from
"http://en.wikipedia.org/wiki/Replicative_transposition
218. General mechanism of
Transposition
Production of protein (enzyme transposase) from the site of
transposase(right corner an Tn 5) should be shown.{the site in upper
diagram in between IR of IS element.}
Action/Motion-Production of protein (enzyme transposase) from the site
of
219. Replicative
Transposition
Single stranded cuts are made on
either side of the Transposon and
on the opposite sides of the
target of the recipient.
220. This produces 4 free ends
get in each DNA molecule
Two of the ends from the donor are
ligated to 2 of the ends of target.
This links the two molecules with a
single molecule of transposon.
221. The two remaining free 3’ ends are
used as primers for DNA polymerase
which uses the Transposon DNA
as the template.This replicates the
transposon and leaves the cointegrate.
228. First, the transposase makes a double-stranded cut in the
donor DNA at the ends of the transposon
and makes a staggered cut
in the recipient DNA.
229. Each end of the donor DNA is then
joined to an overhanging
end of the recipient DNA.
230. DNA polymerase fills in the short,
overhanging sequences,
resulting in a short, direct repeat
on each side of the transposon
insertion in the recipient DNA.
231. INSTRUCTIONS SLIDE
1
Questionnaire to test the user
Q1]Define tranposition?
Transposons sequences DNA
2 the genome ofaresingle cell, ofprocessthat can move around to different positions within
a a called transposition.
Q2]Give examples of non composite transposons.
IS1, IS3, Tn5, Tn10, and Tn3) or a multisubunit complex (e.g. Tn7)
Q3]Describe the general structure of bacterial transposons.
3 Ans
4
5
232. This transposon consists of a
chloramphenicol-resistance gene (dark
blue) flanked by two copies of IS1
(orange), one of the smallest IS elements.
Other copies of IS1, without the drug-
resistance gene, are located elsewhere in
the E. coli chromosome. The internal
inverted repeats of IS1 abutting the
resistance gene are so mutated that
transposase does not recognize them.
During transposition, the IS-element
transposase makes cuts at the positions
indicated by small red arrows, so the
entire transposon is moved from the
donor DNA (e.g., a plasmid). The target-
site sequence at the point of insertion
becomes duplicated on either side of the
233. Q4]Explain the mobile genetic elements found in bacteria.
ANS:-
Three of the many types of mobile genetic elements found in
bacteria. Each of these DNA elements contains a gene that
encodes a transposase, an enzyme that conducts at least some of
the DNA breakage and joining reactions needed for the element to
move. Each mobile element also carries short DNA sequences
(indicated in red) that are recognized only by the transposase
encoded by that element and are necessary for movement of the
element. In addition, two of the three mobile elements shown carry
genes that encode enzymes that inactivate the antibiotics
ampicillin (ampR) and tetracycline (tetR). The transposable
element Tn10, shown in the bottom diagram, is thought to have
evolved from the chance landing of two short mobile elements on
either side of a tetracyclin-resistance gene; the wide use of
tetracycline as an antibiotic has aided the spread of this gene
through bacterial populations. The three mobile elements shown
are all examples of DNA-only transposons
237. Applying Your Knowledge
1. Retrotransposon
2. Transposon
3. Both retrotransposons and transposons
4. Neither retrotransposons nor transposons
Which type of transposable element
• Uses a DNA intermediate for transposition?
• Contains long terminal repeats on its ends?
• Generates direct repeats as a result of
transposition?
• Carries a gene for reverse transcriptase?
• Can insert a copy in a new location while leaving
the old copy at the original site?
238. Effects of
Transposition
Transposable elements can:
Cause mutations in adjacent
genes
Cause chromosomal
rearrangements
Relocate genes
239. Possible Advantages of Transposable
Elements
Transposable elements may:
Create genetic diversity
Act as promoters
Allow recombination between
plasmid and genomic DNA
when multiple copies of the
element are present
Carry antibiotic resistance
genes, conferring an
advantage on bacterial cells
Increase the number of
copies of an exon or gene
240. Examples of Transposable
Elements
Bacterial Insertion Sequences
and
more Complex Transposons
Ac-Ds Elements in Corn
P elements in Fruit Flies
241. Transposable Elements in
Bacteria
Insertion Sequences contain only the
elements needed for transposition
Composite Transposons contain DNA that
has insertion sequences on both sides
Antibiotic
resistance genes
are often included
242. Ac and Ds Elements
in Corn
Ac stands for activator element
Ds stands for dissociative
element
Barbara McClintock showed that
--transposition of the Ds element
altered kernel coloration
--movement of the Ds element
required the activity of Ac
element
245. Relatedness of Ac and Ds
Elements
For transposition, Ds elements require the
transposase produced by the Ac element.
246. Autonomous and Non-
autonomous Elements
Type Properties Example
Autonomous •Can transpose without Ac
the presence of
another element
Non- •Requires the
Ds
autonomous presence of another
functional element
to move
•Autonomous element
provides transposase
or reverse transriptase
247. The P Element in Drosophila Codes for
a Transposase and a Repressor of
Transposition
No
Repressor repressor
produced
P element
Transposition is inserts in
repressed multiple
locations
248. Use of the P Element As a Vector in
Drosophila
P element codes for transposase
P element with gene of interest can insert into chromosomes
with help of plasmid containing only transposase.
249. Applying Your
Knowledge
1. Ac-Ds Elements
2. Alu Element
3. Insertion Sequence
4. P element
Which type of transposable element
• Contains only the sequences needed for
transposition in bacteria?
• Represents a SINE found in humans?
• Is used to insert genes into fruit fly
chromosomes?
• Causes reversible alterations for kernel color
in corn?
Editor's Notes
Left image are sketches of McClintock’s observation of chromosomes in Maize.
The majority of transposition events are ancient, but in Sue Wessler’s story, we se tranpsons in action.