

Recomendados
Mais conteúdo relacionado
Destaque
Destaque (20)
Mais de Victor Soto
Mais de Victor Soto (20)
8 fisiologia esfuerzo-deporte
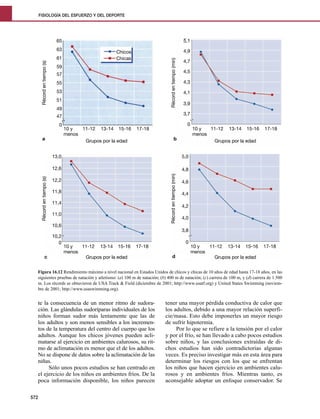
- 1. FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE 65 5,1 63 4,9 Chicos 61 Chicas Récord en tiempo (min) 4,7 Récord en tiempo (s) 59 4,5 57 55 4,3 53 4,1 51 3,9 49 3,7 47 0 0 10 y 11-12 13-14 15-16 17-18 10 y 11-12 13-14 15-16 17-18 menos menos a Grupos por la edad b Grupos por la edad 13,0 5,0 12,6 4,8 Récord en tiempo (min) Récord en tiempo (s) 12,2 4,6 11,8 4,4 11,4 4,2 11,0 4,0 10,6 3,8 10,2 0 0 10 y 11-12 13-14 15-16 17-18 10 y 11-12 13-14 15-16 17-18 menos menos c Grupos por la edad d Grupos por la edad Figura 16.12 Rendimiento máximo a nivel nacional en Estados Unidos de chicos y chicas de 10 años de edad hasta 17-18 años, en las siguientes pruebas de natación y atletismo: (a) 100 m de natación; (b) 400 m de natación; (c) carrera de 100 m, y (d) carrera de 1.500 m. Los récords se obtuvieron de USA Track & Field (diciembre de 2001; http://www.usatf.org) y United States Swimming (noviem- bre de 2001; http://www.usaswimming.org). te la consecuencia de un menor ritmo de sudora- tener una mayor pérdida conductiva de calor que ción. Las glándulas sudoríparas individuales de los los adultos, debido a una mayor relación superfi- niños forman sudor más lentamente que las de cie/masa. Esto debe imponerles un mayor riesgo los adultos y son menos sensibles a los incremen- de sufrir hipotermia. tos de la temperatura del centro del cuerpo que los Por lo que se refiere a la tensión por el calor adultos. Aunque los chicos jóvenes pueden acli- y por el frío, se han llevado a cabo pocos estudios matarse al ejercicio en ambientes calurosos, su rit- sobre niños, y las conclusiones extraídas de di- mo de aclimatación es menor que el de los adultos. chos estudios han sido contradictorias algunas No se dispone de datos sobre la aclimatación de las veces. Es preciso investigar más en esta área para niñas. determinar los riesgos con los que se enfrentan Sólo unos pocos estudios se han centrado en los niños que hacen ejercicio en ambientes calu- el ejercicio de los niños en ambientes fríos. De la rosos y en ambientes fríos. Mientras tanto, es poca información disponible, los niños parecen aconsejable adoptar un enfoque conservador. Se 572
- 2. 02 Cap 07-15 • 221-548 17/1/07 16:31 Página 497 PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE no se establecen adecuadamente, los deportistas M TABLA 14.4 Intervalo de valores para el porcentaje de pueden ser inducidos a reducir su peso muy por grasa corporal de deportistas masculinos y femeninos en debajo del nivel óptimo. Por lo tanto, es de im- varios deportes portancia crítica establecer estándares de peso % de grasa adecuados. Los estándares de peso corporal deben ba- Deporte Hombres Mujeres sarse en la composición corporal del deportista. Una vez determinada la composición corporal, la Béisbol/softball 8-14 12-18 cantidad de masa magra se usa para calcular lo Baloncesto 6-12 10-16 que debe pesar el deportista para un nivel especí- fico de porcentaje de grasa corporal. Considere- Culturismo 5-8 6-12 mos un ejemplo en que el objetivo es hacer que una mujer nadadora de 72,5 kg con un 25% de Piragüismo/kayak 6-12 10-16 grasa corporal descienda hasta el 18%, tal como Ciclismo 5-11 8-15 se muestra en la tabla 14.3. Sabemos que su peso objetivo estará compuesto por un 18% de grasa y Esgrima 8-12 10-16 por un 82% de masa magra. Por lo tanto, para Fútbol americano 6-18 –– calcular su peso objetivo con un 18% de grasa corporal, dividiremos su masa magra por el 82%, Golf 10-16 12-20 que es la fracción de su peso objetivo que tiene que Gimnasia 5-12 8-16 estar representado por su masa magra. Este cálculo (54,4 kg dividido por 0,82) nos da un peso objeti- Hípica 6-12 10-16 vo de 66,3 kg, por lo que esta mujer necesita per- der 6,2 kg. Hockey sobre hielo/hierba 8-16 12-18 Por lo tanto, el establecimiento de estándares Orientación 5-12 8-16 de peso debe sustituirse por el de estándares de porcentaje de grasa corporal para cada deporte. Pentatlón ––– 8-15 Teniendo esto presente, ¿cuál es el porcentaje de Frontón con raqueta 6-14 10-18 grasa corporal recomendado para un deportista de elite en un deporte dado? Para cada deporte Remo 6-14 8-16 debe establecerse un valor o un intervalo de valo- res óptimos de grasa relativa fuera de los cuales Rugbi 6-16 –– es probable que el rendimiento del deportista se Patinaje sobre hielo 5-12 8-16 deteriore. Y puesto que la distribución de la grasa Esquí 7-15 10-18 Saltos de esquí 7-15 10-18 M TABLA 14.3 Cálculo del peso objetivo para la obten- Fútbol 6-14 10-18 ción del rendimiento óptimo en una nadadora Natación 6-12 10-18 Peso 72,5 kg Natación sincronizada –– 10-18 Grasa relativa 25% Tenis 6-14 10-20 Peso de grasa 18,2 (72,5 kg × 0,25%) Pista y campo Peso magro 54,4 kg (72,5 kg - 18 kg de peso Pruebas de carrera 5-12 8-15 de grasa) Pruebas de campo 8-18 12-20 % objetivo de grasa 18% (= 82% de masa magra) Triatlón 5-12 8-15 Balonvolea 7-15 10-18 Peso objetivo 66,3 kg (= 54,4 kg/0,82) Levantamiento de pesos 5-12 10-18 Objetivo de pérdida de peso 6,35 kg Lucha libre 5-16 –– 497
- 3. 02 Cap 07-15 • 221-548 17/1/07 16:29 Página 312 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE Ventilación pulmonar i El volumen sanguíneo aumenta como Después del entrenamiento, la ventilación pul- consecuencia del entrenamiento de fon- monar permanece esencialmente invariable o se do. reduce levemente en reposo, y disminuye ligera- mente a ritmos de esfuerzo submáximos estanda- i El incremento está ocasionado princi- rizados. Pero la ventilación pulmonar máxima palmente por un aumento en el volumen aumenta sustancialmente. Normalmente, los in- de plasma sanguíneo. crementos en sujetos no entrenados son desde un ritmo inicial de 120 l/min hasta un ritmo de apro- i El número de glóbulos rojos puede au- ximadamente 150 l/min después del entrena- mentar, pero la ganancia de plasma es miento. Los ritmos de ventilación pulmonar nor- normalmente mucho más elevada, lo malmente aumentan hasta unos 180 l/min en que da lugar a una porción fluida de la deportistas muy entrenados. Dos son los factores sangre relativamente mayor. que pueden ser responsables del incremento en la ventilación pulmonar máxima después del entre- i El aumento del volumen plasmático incre- namiento: el mayor volumen oscilante y la mayor menta la viscosidad de la sangre, lo cual frecuencia respiratoria en el ejercicio máximo. mejora la circulación y la disponibilidad Los estudios recientes revelan que el entrena- de oxígeno. miento de los músculos inspiratorios puede me- jorar significativamente el rendimiento. En un es- i Determinadas investigaciones han de- tudio con 14 remeras competitivas, 11 semanas mostrado que los cambios en el volumen de entrenamiento de los músculos inspiratorios de plasma guardan una estrecha relación mejoraron la distancia recorrida en un esfuerzo con los producidos en el volumen sistó- máximo durante 9,6 kilómetros remando (3,5%) lico y en el nVO2 máx, lo que provoca y en la marca para completar una contrarreloj de que el incremento del volumen de plas- 5.000 m (3,1%).46 ma inducido por el ejercicio sea uno de los efectos más significativos del entre- namiento. Los grandes deportistas con un alto nivel de entrenamiento de fondo, tales como los remeros, pueden tener índices de ven- tilación pulmonar máxima de 240 l/min, más del doble del ritmo normal de indivi- Adaptaciones respiratorias duos no entrenados. al entrenamiento Independientemente de lo eficaz que sea el siste- ma cardiovascular suministrando cantidades Generalmente, la ventilación no se considera adecuadas de sangre a los tejidos, la capacidad un factor limitante del rendimiento en los ejer- de resistencia se verá perjudicada si el sistema cicios de resistencia. No obstante, existen prue- respiratorio no aporta suficiente oxígeno para sa- bas que indican que, en un determinado momento tisfacer las demandas. El funcionamiento del sis- de la adaptación de una persona muy entrenada, tema respiratorio no suele limitar el rendimiento la capacidad del sistema pulmonar para transpor- porque la ventilación puede incrementarse en tar oxígeno no podrá satisfacer las demandas de mayor medida que la función cardiovascular. Pe- las extremidades y del sistema cardiovascular.9 ro, al igual que con el sistema cardiovascular, el Esto causa lo que se ha denominado hipoxemia sistema respiratorio experimenta adaptaciones arterial inducida por el ejercicio, en la que la sa- específicas al entrenamiento de fondo para ma- turación arterial de oxígeno disminuye por deba- ximizar su eficacia. Consideremos algunas de jo del 96%. La hipoxemia arterial leve inducida ellas. por el ejercicio se define como una saturación ar- terial del 93% al 95%, moderada del 88% al 93% y grave de menos del 88%.10 312
- 4. 02 Cap 07-15 • 221-548 17/1/07 16:32 Página 533 AYUDAS ERGOGÉNICAS Y RENDIMIENTO de, nuestra resistencia aeróbica y, por lo tanto, las controversias que originaban los primeros es- nuestro rendimiento, puede incrementarse sus- tudios. Muchos de los estudios iniciales que no tancialmente. Ésta es la premisa que subyace ba- hallaron ninguna mejora con el dopaje sanguíneo jo el dopaje sanguíneo. habían reinyectado sólo pequeños volúmenes de glóbulos rojos y la reinfusión se efectuó antes de pasadas 3 ó 4 semanas de la extracción de sangre. Efectos demostrados Primero, parece ser que hay que extraer y rein- Ekblom y colaboradores27 provocaron una con- yectar 900 ml de sangre, o incluso más cantidad. moción en el mundo deportivo a comienzos de la Los incrementos en el nVO2 máx y en el rendi- década de 1970. En uno de sus estudios, retiraron miento no son tan grandes cuando se usan volú- entre 800 y 1.200 ml de sangre de sus sujetos y menes más pequeños. De hecho, algunos estudios luego volvieron a inyectarles los glóbulos rojos que usan volúmenes pequeños no han logrado ha- unas 4 semanas más tarde. Los resultados mos- llar ninguna diferencia. traron una mejora considerable en el nVO2 máx Segundo, parece que hay que esperar por lo (9%) y en el tiempo en la cinta ergométrica menos de 5 a 6 semanas y posiblemente hasta 10 (23%). Inmediatamente después, durante unos semanas antes de la reinfusión. Esto se basa en el pocos años, varios estudios no lograron confir- tiempo requerido por el cuerpo para restablecer su mar estos originales descubrimientos, pero otros hematocrito previo a la extracción de sangre. varios mostraron un efecto ergogénico. Por último, estudios iniciales refrigeraron la Por lo tanto, la literatura de las investigacio- sangre retirada. El tiempo máximo de almacenaje nes estuvo dividida sobre la eficacia del dopaje bajo refrigeración es de, aproximadamente, 5 se- sanguíneo hasta que en 1980 se produjo un im- manas. Además, cuando la sangre es refrigerada, portante adelanto como consecuencia de un estu- aproximadamente el 40% de los glóbulos rojos se dio realizado por Buick y colaboradores.13 Se destruyen o se pierden. Estudios posteriores han examinó a 11 corredores de fondo muy entrena- usado la congelación para almacenar la sangre. La dos en diferentes momentos durante el estudio: congelación permite un tiempo de almacenaje ca- si ilimitado y solamente se pierden aproximada- (1) antes de la extracción de sangre; (2) des- mente el 15% de los glóbulos rojos. pués de dar al cuerpo un tiempo apropiado para Gledhill30, 31 concluyó que el dopaje sanguí- restablecer los niveles normales de glóbulos ro- neo produce mejoras significativas en el nVO2 jos, pero antes de la reinfusión de la sangre extraí- máx y en la capacidad de resistencia cuando se da; (3) después de una simulada reinfusión de 50 hace bajo condiciones óptimas: ml de una solución salina (un placebo); (4) des- • Un mínimo de 900 ml de reinfusión de san- pués de la reinfusión de 900 ml de sangre que ori- gre. ginalmente se habían extraído y preservado conge- • Un intervalo mínimo de 5 a 6 semanas en- lándolos; (5) después de que los niveles elevados tre la extracción y la reinfusión. de glóbulos rojos hubiesen vuelto a la normali- • Almacenaje de la sangre por congelación. dad. Tal como se muestra en la figura 15.5, los in- Gledhill ha demostrado también que estas vestigadores hallaron un incremento sustancial mejoras son la consecuencia directa del mayor en el nVO2 máx y en el tiempo en la cinta ergomé- contenido de hemoglobina de la sangre, no del trica después de la reinfusión de los glóbulos ro- mayor gasto cardíaco debido a un incremento del jos (grupo A) y ningún cambio después de la fal- volumen del plasma. sa reinfusión (grupo B). Este incremento en el nVO2 máx persistió durante un máximo de 16 se- Dopaje sanguíneo y capacidad de resistencia manas, pero los incrementos del tiempo en la cin- ta ergométrica se redujeron antes de transcurridos ¿Produce el incremento del nVO2 máx y del tiempo los primeros 7 días. en la cinta ergométrica, como consecuencia del dopaje sanguíneo, una mejora en la capacidad de resistencia? Diversos estudios han analizado esta Maximización de los beneficios cuestión. En un estudio se observaron tiempos de ¿Por qué fue un gran adelanto el estudio de Buick carrera de 8 km sobre cinta ergométrica en un gru- y colaboradores? Gledhill30, 31 ayudó a explicar po de 12 corredores de fondo experimentados.63 533
- 5. 02 Cap 07-15 • 221-548 17/1/07 16:30 Página 392 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE Músculos Estudios sobre la inmovilización han mostrado la existencia de cambios muy rápidos en la estruc- tura y en la función de los músculos cuando las extremidades se hallan inmovilizadas con yeso o % de cambio cuando las extremidades posteriores de los ani- males cuadrúpedos están suspendidas. La atrofia muscular es la consecuencia principalmente de una menor síntesis de proteínas.32 Experimentos con ratas demostraron que la síntesis de proteínas disminuye aproximadamente en un 35% durante 28 días en el laboratorio las primeras horas y hasta en un 50% en los pri- espacial (n = 3) 30 días de reposo meros días de inmovilización, lo cual conduce a en cama (n = 8) una pérdida neta de proteína muscular.3, 40 Con el tiempo, la atrofia muscular resultante puede lle- Fuerza Fuerza Volumen Peso gar a ser importante. Estudios parecidos con hu- de los de los de las corporal manos han demostrado una pérdida más lenta de músculos músculos piernas masa muscular y proteínas durante la anulación flexores de extensores la rodilla de la rodilla de peso en carga sobre los músculos (es decir, re- poso en cama y suspensión de las piernas).42,50 Asimismo, existe una importante diferencia entre la inmovilización y la microgravedad. Con la in- % de cambio movilización, los músculos experimentan poca o ninguna activación. En cambio, con la microgra- vedad los músculos se activan, pero se cargan con una intensidad menor debido a la pérdida de De contracción lenta los efectos de la gravedad. Durante una misión en De contracción el espacio de 17 días en 1996 (STS-78), pocos o rápida ningún cambio se obtuvieron en la fuerza de los músculos de la pantorrilla de los cuatro miem- Vasto lateral Sóleo bros de la tripulación a pesar de que las fibras musculares perdieron entre un 8% y un 11% del Figura 11.11(a) Fuerza y volumen de piernas, y cambios en el pe- área transversal.14 so corporal con 30 días de reposo en cama y un vuelo espacial de Estudios llevados a cabo sobre sujetos que 28 días. (b) Cambios en el área transversal de las fibras muscula- guardan reposo en cama, simulando la microgra- res de contracción lenta y rápida tras 30 días de reposo en cama. vedad, han hallado reducciones importantes, tan- Adaptado con autorización de V.A. Convertino, 1991, «Neuro- to de la fuerza como de la sección transversal, de muscular aspects in development of exercise countermeasures», las fibras musculares ST y FT, con tendencia a The Physiologist 34: S125-S128. una mayor afectación de las fibras FT.10 En la fi- gura 11.11 se ilustran estas relaciones; obsérvese la estrecha coincidencia de los cambios en la fuerza y en el volumen de las piernas, así como duración de las misiones, por el programa de del peso corporal entre un ensayo de reposo en ejercicios en vuelo, por diferencias en los grupos cama de 30 días de duración para simular la mi- de músculos, o por una combinación de estos crogravedad y el vuelo de 28 días de duración tres factores. del Skylab 2. A diferencia de los músculos de la pantorrilla en la misión espacial de 17 días de los que hemos hablado en el párrafo anterior, los músculos del muslo durante esta misión de 28 La fuerza y la sección transversal de las días en el laboratorio espacial perdieron fuerza, fibras musculares ST y FT se reducen hasta un 20% en el caso de los músculos exten- con la exposición a la microgravedad si- sores de la rodilla. La diferencia en los resulta- mulada y a la microgravedad real. dos de estos dos vuelos podría explicarse por la 392
- 6. 02 Cap 07-15 • 221-548 17/1/07 16:29 Página 311 ADAPTACIONES CARDIOVASCULARES Y RESPIRATORIAS AL ENTRENAMIENTO hematíes a lo largo de años de entrenamiento de- Volumen de plasma, volumen vuelve la relación entre la masa celular y el volu- sistólico y nVO2 máx men total a niveles más normales. Este cambio en la proporción entre plasma y Un incremento del volumen de plasma es uno de células, resultante de un incremento en la porción los cambios más significativos que se producen fluida, reduce la viscosidad de la sangre. Ello con el entrenamiento de fondo. Recordemos de puede facilitar el movimiento de la sangre por los nuestra anterior discusión que el incremento del vasos sanguíneos, particularmente a través de los volumen de plasma es un factor importante en el vasos más pequeños, como son los capilares. De- aumento del volumen sistólico resultante del en- terminadas investigaciones han demostrado que trenamiento. El volumen sistólico, a su vez, afec- la baja viscosidad de la sangre aumenta el aporte ta al consumo de oxígeno. de oxígeno a las masas musculares activas. Cuando el volumen de plasma aumenta, tam- Tanto la cantidad total de hemoglobina (valo- bién lo hace el volumen sanguíneo. En conse- res absolutos) como el número total de glóbulos cuencia, entra más sangre en el corazón. Cuando rojos en deportistas altamente entrenados general- entra más sangre en el corazón, el volumen sistó- mente son superiores a lo normal, aun cuando los lico aumenta. A intensidades máximas de esfuer- valores relativos son inferiores a los normales. zo, el nVO2 máx suele permanecer relativamente Ello asegura que la sangre tenga una capacidad de estable, por lo que un mayor volumen sistólico transporte de oxígeno más que suficiente para sa- permite que el gasto cardíaco máximo se incre- tisfacer las necesidades del cuerpo en todo mo- mente. El aumento del gasto cardíaco máximo mento. El índice del recambio metabólico de he- hace que haya más oxígeno disponible para los matíes también podría ser mayor con un músculos activos, permitiendo así que aumente el entrenamiento intenso. Esto tal vez sea una venta- nVO2 máx. ja, siempre que la producción sea igual o mayor que la destrucción, dado que las células jóvenes son más eficaces en el transporte de oxígeno.42 El incremento del volumen sanguíneo Volumen Volumen con el entrenamiento aeróbico de fondo sanguíneo = 5 l sanguíneo = 5,7 l se debe al gran aumento del volumen Hematocrito = 44% Hematocrito = 42% plasmático y a un pequeño incremento del número de hematíes; ambos cam- bios facilitan la llegada de oxígeno a los músculos activos. Esta secuencia de acontecimientos –un au- Volumen (l) 2,8 3,3 mento del volumen de plasma que conlleva un Volumen Volumen aumento del volumen sistólico, que, a su vez, plasmático plasmático conduce a un aumento en el nVO2 máx– se ha de- mostrado que también tiene lugar en individuos no entrenados cuyos volúmenes de plasma se 2,2 2,4 han incrementado usando una infusión de dex- Hematíes Hematíes trano al 6%.24 Esta secuencia también funciona a la inversa: una reducción del volumen de plasma por el abandono de los entrenamientos por parte Preentrenamiento Posentrenamiento de deportistas muy entrenados, o situando a hombres sanos en una posición de tendido con la cabeza hacia abajo (–5°) durante 20 h se obtie- Figura 9.10 Aumento en la volemia total y el volumen plasmá- nen reducciones significativas en el volumen sis- tico con un entrenamiento de fondo. Hay que reparar en que, aunque el hematocrito (porcentaje de hematíes) disminuye del tólico y disminuciones equivalentes en el nVO2 44% al 42%, el volumen total de hematíes aumenta un 10%. máx.8, 18 311
- 7. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 126
- 8. 02 Cap 07-15 • 221-548 17/1/07 16:31 Página 499 PESO CORPORAL, COMPOSICIÓN CORPORAL Y DEPORTE Evitar el ayuno y las dietas de choque La figura 14.8 ilustra estos cambios en el agua, la grasa, las proteínas y los hidratos de car- Nuestro jugador de fútbol americano debe perder bono del cuerpo durante un período de 30 días de unos 2 kg por semana durante las 4 semanas si- ayuno. El agotamiento de los hidratos de carbono guientes, por lo que decide embarcarse en una se produce antes de transcurridos 3 días. El con- dieta de choque, y elige cualquier dieta de las que tenido de proteínas cae significativamente en el están de moda, sabiendo que una persona puede mismo período de tiempo. El contenido de agua perder unos 3 kg por semana con tales dietas. No del cuerpo disminuye abruptamente durante los es un caso aislado. Muchos deportistas descubren primeros 3 días. El ritmo de pérdida de grasa, que tienen un peso excesivo y que no están en aunque disminuye algo en el segundo y tercer forma porque han comido en exceso y reducido días, permanece relativamente constante durante su actividad al finalizar la temporada y suelen es- el resto del período de ayuno. perar hasta las últimas semanas antes de la fecha Para la mayoría de la gente es imposible en que deben presentarse para atajar el problema. perder más de 2 kg de grasa por semana, incluso En nuestro ejemplo, el jugador de fútbol america- en condiciones de ayuno completo. Esto se pue- no puede ser capaz de perder 9 kg en 4 semanas de demostrar fácilmente. Si 0,5 kg de grasa (teji- con la dieta de choque. Pero una gran parte de es- do adiposo) tienen un equivalente calórico de ta pérdida de peso lo será de agua corporal y muy 3.500 kcal, se necesita un déficit de 3.500 kcal poca de grasa acumulada. Varios estudios han in- para perder 0,5 kg de grasa. El ritmo metabólico formado de que se producen sustanciales pérdidas en reposo del jugador profesional de fútbol ame- de peso con dietas muy bajas en calorías (500 kcal por día o menos); pero de la pérdida de peso, más del 60% procede de tejidos magros y menos del 40% de depósitos de grasa. Días de ayuno Aunque una gran parte del peso de los juga- 0 3 5 10 15 20 25 30 dores de fútbol americano se pierde disminuyen- 0 do las reservas de agua, también se pierde una sustancial cantidad de proteínas. Asimismo, la 100 mayoría de las dietas de choque se basan en una 250 importante reducción de la ingestión de hidratos de carbono. Esta menor ingestión es insuficiente para abastecer las necesidades del cuerpo en 500 cuanto a hidratos de carbono, y en consecuencia las reservas de estos se agotan. Puesto que la acu- Pérdida de peso (g/24 h) mulación de agua acompaña el almacenaje de hi- dratos de carbono, las reservas de agua también se reducen cuando disminuyen las reservas de aquellos. Por cada gramo de hidratos de carbono que emplea el cuerpo, se pierden aproximada- 1.000 mente 2,6 g de agua. El contenido total de glucó- Hidratos de carbono geno del cuerpo es de 800 g a 1.000 g, por lo que Proteínas la depleción del glucógeno almacenado provoca- Grasas ría una pérdida aproximada de 2.080-2.600 g de agua, en torno a 2-2,6 kg. Agua Además, el cuerpo depende en mayor medi- 1.500 da de los ácidos grasos libres para obtener ener- gía cuando se agotan las reservas de hidratos de carbono. En consecuencia, los cuerpos cetónicos, un subproducto del metabolismo de los ácidos grasos, se acumulan en la sangre y producen una condición conocida como cetosis. Esta condición 2.000 incrementa todavía más la pérdida de agua. Una gran parte de esta pérdida de agua se produce du- Figura 14.8 Composición de la pérdida de peso con 30 días de rante la primera semana de dieta. ayuno. 499
- 9. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 99 ADAPTACIONES NEUROMUSCULARES AL ENTRENAMIENTO RESISTIDO mizar la fuerza generada por un músculo agonis- trovertidas. Sin embargo, es de importancia el re- ta, es necesario reducir al mínimo el grado de co- ciente descubrimiento que indica que el compo- activación.14 La reducción en la coactivación po- nente excéntrico del entrenamiento es importante dría explicar una porción de aumento de la fuerza para los incrementos máximos en la sección atribuida a factores neuronales, pero su contribu- transversal de la fibra muscular.14, 30 En estudios ción sería pequeña. llevados a cabo con sujetos que realizaban sólo La codificación del índice es otro factor acciones concéntricas o excéntricas, se observó, potencial que puede aumentar la producción después de 36 sesiones de entrenamiento, que el de fuerza de un músculo después del entrena- entrenamiento excéntrico producía un aumento miento. La codificación del índice es un térmi- en la fibra FT y de la fuerza aproximadamente 10 no empleado para describir la frecuencia de veces mayor que el alcanzado con el entrena- activación o la frecuencia de descarga de uni- miento concéntrico. Entonces, el entrenamiento dades motoras. Aunque no se haya estudiado sólo con acciones concéntricas podría limitar al bien, hay ciertas pruebas que lo respaldan co- músculo hipertrofiado. Echemos un vistazo a mo un factor posible. 17 También se han apre- dos mecanismos de los que se ha postulado que ciado cambios en la morfología de la unión aumentan el tamaño muscular con el entrenamien- neuromuscular con el aumento y reducción de to resistido: la hipertrofia y la hiperplasia. los niveles activos que tal vez estén directa- mente relacionados con la capacidad de pro- ducción de fuerza del músculo. 11 Hipertrofia de las fibras Ahora que hemos examinado los factores Las primeras investigaciones indicaban que el neurológicos potenciales que contribuyen a au- número de fibras musculares de cada uno de mentar la fuerza con entrenamiento resistido, pres- nuestros músculos queda establecido en el naci- temos atención a la hipertrofia muscular. Estu- miento o poco después, y que este número per- diaremos la forma en que los músculos aumentan manece invariable a lo largo de la vida. Si esto es de tamaño y cómo ello contribuye al incremento de cierto, entonces la hipertrofia crónica sólo puede la fuerza, o a la capacidad de producción de fuerza ser la consecuencia de la hipertrofia de fibras del músculo. musculares individuales. La explicación de ello puede radicar en: Hipertrofia muscular • un mayor número de miofibrillas; ¿Cómo aumenta el tamaño de un músculo? Dos tipos de hipertrofia pueden tener lugar: temporal • más filamentos de actina y de miosina; y crónica. La hipertrofia temporal es este abul- • más sarcoplasma; tamiento del músculo que tiene lugar durante una sola serie de ejercicios. Es el resultado principal- • más tejido conectivo, o mente de la acumulación de fluidos (edema) en • cualquier combinación de lo anterior. los espacios intersticiales e intracelulares del músculo. Este fluido lo pierde el plasma sanguí- neo.42 La hipertrofia temporal, como su nombre Tal como se observa en las micrografías de indica, dura tan sólo un corto período de tiempo. la figura 3.4, el entrenamiento intenso contra re- El fluido vuelve a la sangre al cabo de unas horas sistencia puede incrementar notablemente la sec- de haber finalizado el ejercicio. ción transversal de las fibras musculares. En este La hipertrofia crónica se refiere al incre- ejemplo, la hipertrofia de las fibras es producida mento en el tamaño muscular que se produce me- probablemente por un mayor número de miofibri- diante el entrenamiento resistido a largo plazo. llas y de filamentos de actina y de miosina, que Esto refleja verdaderos cambios estructurales en proporcionarán más puentes cruzados para la el músculo como resultado de un aumento del producción de fuerza durante la contracción má- número de fibras musculares (hiperplasia) o co- xima. Estos espectaculares agrandamientos de las mo consecuencia de un aumento del tamaño de fibras musculares no ocurren, sin embargo, en to- las fibras musculares individuales existentes (hi- dos los casos de hipertrofia muscular. pertrofia). Las teorías que intentan explicar la La hipertrofia de las fibras individuales me- causa subyacente de este fenómeno son muy con- diante entrenamiento resistido parece ser pro- 99
- 10. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:21 Página 131 METABOLISMO, ENERGÍA Y SISTEMAS BÁSICOS DE ENERGÍA ye nuestro cuerpo, y proporcionan generalmente poca energía para la función celular. Al pasar de Las reservas de hidratos de carbono en el la realización de un esfuerzo muscular suave a hígado y en los músculos esqueléticos están otro más duro, se emplean progresivamente más limitadas a menos de 2.000 kcal de energía, hidratos de carbono, dependiendo menos de las o el equivalente de la energía necesaria grasas. En los ejercicios máximos de corta dura- para correr aproximadamente 32 km. ción, el ATP se genera casi exclusivamente a par- Las reservas de grasa, no obstante, sue- tir de los hidratos de carbono. len superar las 70.000 kcal de energía acumulada. Hidratos de carbono La dependencia de nuestros músculos respecto a los hidratos de carbono durante el ejercicio está Tal como se ve en la figura 4.1, se obtiene relacionada con la disponibilidad de hidratos de sustancialmente más energía de una cantidad de- carbono y con que el sistema muscular esté bien terminada de grasa (9 kcal/g) que de la misma desarrollado para su metabolismo. Los hidratos cantidad de hidratos de carbono (4 kcal/g). No de carbono se convierten en última instancia en obstante, el ritmo de liberación de energía de es- glucosa, un monosacárido (azúcar de una sola tos compuestos es demasiado lento para satisfa- unidad) que es transportado por la sangre a los te- cer todas las demandas de energía de la actividad jidos activos, donde se metaboliza. En reposo, la muscular intensa. ingesta de hidratos de carbono es absorbida por los músculos y el hígado, y luego se convierte en una molécula de azúcar mucho más compleja: Proteínas glucógeno. Éste se almacena en el citoplasma hasta que las células lo utilizan para formar ATP. El proceso por el que las proteínas o las grasas se El glucógeno, depositado en el hígado, cuando se convierten en glucosa recibe el nombre de gluco- necesita, se convierte nuevamente en glucosa que neogénesis. Alternativamente, las proteínas pue- es transportada por la sangre a los tejidos activos den convertirse, a través de una serie de reaccio- donde es metabolizada. nes, en ácidos grasos. Esto recibe el nombre de Las reservas de glucógeno en el hígado y en lipogénesis. los músculos son limitadas y pueden agotarse a Las proteínas pueden aportar entre el 5% y el menos que la dieta contenga una razonable canti- 10% de la energía necesaria para mantener un dad de hidratos de carbono. Por lo tanto, depen- ejercicio prolongado. Sólo las unidades más bási- demos fuertemente de nuestras fuentes dietéticas cas de las proteínas (los aminoácidos) pueden de almidones y azúcares para reponer nuestras usarse para obtener energía. Un gramo de proteí- reservas de hidratos de carbono. Sin una inges- na genera alrededor de 4,1 kcal. tión adecuada de ellos, los músculos y el hígado pueden quedar desprovistos de su principal fuen- te de energía. Ritmo de liberación de energía Para ser útil, la energía libre debe liberarse a partir de compuestos químicos a un ritmo controlado. Grasas Este ritmo viene parcialmente determinado por la Las grasas y las proteínas también se usan como elección de la fuente primaria de combustible. fuentes energéticas. Nuestro cuerpo acumula mu- Grandes cantidades de un combustible determina- cha más grasa que hidratos de carbono. Tal como do pueden hacer que las células dependan más de se ve en la tabla 4.1, las reservas energéticas del esta fuente que de otras alternativas. Esta influen- cuerpo en grasas es mucho mayor que las de hi- cia de la disponibilidad de energía recibe la deno- dratos de carbono. Pero las grasas son menos ac- minación de efecto de acción de masa. cesibles para el metabolismo celular, porque pri- Enzimas específicas proporcionan un mayor mero deben ser reducidas desde su forma control estructurado del ritmo de liberación de compleja (triglicéridos) a sus componentes bási- energía libre. Muchas de estas proteínas especia- cos: glicerol y ácidos grasos libres. Sólo estos se les facilitan la descomposición (catabolismo) de usan para formar ATP. los compuestos químicos. El camino por el que 131
- 11. 02 Cap 07-15 • 221-548 17/1/07 16:29 Página 335 REGULACIÓN TÉRMICA Y EJERCICIO temperaturas corporales se desvíen fuera de su nemos ahora los mecanismos mediante los cuales intervalo normal de variación de entre 36,1 y se transfiere el calor corporal al ambiente. 37,8 oC. La temperatura corporal refleja la existencia de un cuidadoso equilibrio entre la producción y Transferencia del calor corporal la pérdida de calor. Siempre que este equilibrio se Para que el cuerpo pueda transferir calor al am- altera, la temperatura de nuestro cuerpo cambia. biente, el calor corporal debe poder acceder al Recordemos del capítulo 4 que una gran parte de exterior. El calor de las partes profundas del cuer- la energía que genera nuestro cuerpo se degrada a po (el núcleo) es transportado por la sangre hacia calor, la forma más baja de energía. Todos los teji- la piel (la cáscara). Una vez que el calor se acerca dos metabólicamente activos producen calor que a la piel, puede ser transferido al ambiente por puede usarse para mantener la temperatura interna cualquiera de los cuatro mecanismos siguientes: corporal. Pero si la producción de calor corporal conducción, convección, radiación y evapora- supera la pérdida de calor del cuerpo, nuestra ción. Estos se ilustran en las figuras 10.1 y 10.2. temperatura interna se eleva. Nuestra capacidad para mantener una temperatura interna constante depende de nuestra capacidad para equilibrar el Conducción y convección calor que obtenemos del metabolismo y del am- biente con el calor que pierde nuestro cuerpo. En La conducción del calor supone la transferencia la figura 10.1 se muestra este equilibrio. Exami- de éste desde un material a otro a través del con- Radiación Evaporación del sudor Convección Conducción Glándula sudorípara Calor producido en Calor en la los músculos sangre Figura 10.2 Eliminación de calor por la piel. El calor se dirige a la superficie del cuerpo a través de la sangre arterial y también por con- ducción a través del tejido subcutáneo. Cuando la temperatura de la piel es mayor que la del ambiente, el calor se elimina por conducción, convección, radiación y evaporación del sudor. 335
- 12. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:20 Página 102 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE Un estudio indicó que las áreas medias de fi- • Estudiantes femeninas de educación física: bras musculares de los músculos vasto externo y 4.400 µm2. deltoides eran menores en un grupo de culturistas de alto nivel que en un grupo de referencia de Las diferencias entre estos estudios pueden halterófilos de competición, y eran casi idénticas explicarse por la naturaleza de la carga del entre- a las de estudiantes de educación física y a las de namiento o del estímulo. Se cree que el entrena- personas que no seguían programas de entrena- miento de fuerza. Esto indica que la hipertrofia miento con intensidades elevadas o con grandes de fibras musculares individuales no tenía una resistencias produce una mayor hipertrofia de las importancia crítica para las ganancias en masa fibras, especialmente de las fibras FT, que el en- muscular de los culturistas.57 trenamiento con intensidades o resistencias más Se obtuvieron resultados similares en un es- bajas.35 tudio posterior que comparaba culturistas muy Sólo ha habido un estudio longitudinal que entrenados y controles activos, pero no entrena- haya demostrado la posibilidad de hiperplasia en dos. Las secciones de fibra muscular de los suje- hombres sometidos previamente a entrenamiento tos entrenados eran similares a las de los sujetos resistido a nivel recreativo.39 Después de 12 se- de control, a pesar del hecho de que los primeros manas de entrenamiento intensificado contra re- tenían circunferencias mucho mayores en las ex- sistencia, pareció haber un aumento significativo tremidades.38 Los investigadores también encon- del número de fibras musculares en el bíceps bra- traron más fibras musculares por unidad motora quial de varias de las 12 personas. De este estu- en los culturistas entrenados que en el grupo de dio se deduce que la hiperplasia puede producirse control no deportista. Dado que estos hombres te- en seres humanos, pero sólo en algunas personas nían una circunferencia muscular sustancialmen- y en ciertas condiciones de entrenamiento. te mayor, pero una sección transversal normal de De la información precedente, parece ser fibras musculares, estos descubrimientos indican que la hiperplasia puede darse en animales y po- que el número de fibras musculares aumentó. siblemente en humanos. ¿Cómo se forman estas Una explicación alternativa es que estos deportis- células nuevas? Como se ve en la figura 3.6, se tas tenían un mayor número de fibras al nacer. postula que las fibras musculares individuales tienen capacidad para dividirse en dos células, cada una de las cuales se desarrolla en una fibra muscular funcional. Más recientemente se ha es- tablecido que es probable que las células satéli- Ha quedado ampliamente demostrado tes, que son células madre miógenas implicadas que la hiperplasia de las fibras muscula- en la regeneración del músculo esquelético, inter- res se produce en modelos animales vengan en la generación de nuevas fibras muscu- usando distintos medios para inducir la lares. Estas células suelen activarse después de hipertrofia muscular. Por otra parte, sólo lesiones musculares y, como veremos más ade- unos pocos estudios aportan evidencias lante en este capítulo, las lesiones musculares son de hiperplasia en seres humanos. producto de un entrenamiento intenso, sobre todo el entrenamiento de acciones excéntricas.3 Las le- siones musculares pueden causar una cascada de respuestas, en la que las células satélites se acti- Por otro lado, al menos un estudio ha encon- van y proliferan, migran a la región dañada, y se trado grandes diferencias en la sección de las fi- fusionan con las miofibrillas existentes, o se com- bras musculares al comparar a culturistas con estu- binan y fusionan para producir nuevas fibras.31 diantes de educación física masculinos y Esto se muestra en la figura 3.7. femeninos.44 Las secciones medias de fibra del vasto externo para cada uno de los tres grupos eran como sigue: Integración de la activación neuronal y la hipertrofia fibrilar • Culturistas: 8.400 µm2. Los estudios realizados hasta la fecha sobre las • Estudiantes masculinos de educación físi- adaptaciones del entrenamiento resistido mues- ca: 6.200 µm2. tran que los incrementos iniciales en la fuerza vo- 102
- 13. 02 Cap 07-15 • 221-548 17/1/07 16:31 Página 470 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE reducción del glucógeno muscular y hepático ge- neralmente produce un cambio en el peso corpo- i De un estudio sobre corredores de am- ral de 0,45 a 1,36 kg. Algunos científicos han bos sexos, se dedujo que la mayoría se- propuesto que las reservas de glucógeno muscu- guía una alimentación sana, que cubría lar y del hígado pueden controlarse registrando el la RDR de la mayoría de los nutrientes, peso del deportista a primeras horas de la mañana pero su composición era distinta de la inmediatamente después de levantarse, después esperada, con un 50% del total de las ca- de haber vaciado la vejiga y antes de tomar el de- lorías procedente de hidratos de carbo- sayuno. Una súbita reducción de peso puede re- no, un 36% de grasas, y un 14% de pro- flejar una incapacidad para reemplazar el glucó- teínas. geno, un déficit en el agua corporal, o ambas cosas. i Muchos deportistas han adoptado dietas Los deportistas que deben entrenarse o com- vegetarianas y parecen rendir bien. Sin petir en pruebas agotadoras durante días sucesi- embargo, debe tenerse mucho cuidado vos deben reemplazar las reservas de glucógeno con las fuentes proteicas y el consumo muscular y hepático con la mayor rapidez posi- de niveles adecuados de hierro, zinc, ble. Aunque el glucógeno hepático puede agotar- calcio y vitaminas. se totalmente al cabo de 2 horas de ejercicio al 70% del nVO2 máx, se recupera antes de transcu- i La comida de precompetición debería rridas unas pocas horas cuando se toma una dieta consumirse no antes de las 2 horas pre- rica en hidratos de carbono. Por otro lado, la re- vias a la competición, y debería ser baja síntesis del glucógeno muscular es un proceso en grasas y fácilmente digerible. Una lento, que precisa varios días para volver a la nor- comida líquida de precompetición baja malidad después de una sesión de ejercicio ago- en grasas y rica en hidratos de carbono tador como puede ser el maratón (figura 13.10).7, presenta ventajas. 50 Estudios llevados a cabo a finales de los años 1980 revelaron que la resíntesis del glucógeno i La carga de hidratos de carbono aumen- muscular alcanzaba su mayor velocidad cuando ta en gran medida el contenido de glucó- los individuos tomaban al menos 50 g (aproxima- geno de los músculos, lo cual, a su vez, damente, 0,7 g por kg de peso corporal) de gluco- potencia el rendimiento de fondo. sa cada 2 horas después del ejercicio.8, 34 Tomar cantidades superiores a éstas no parece que ace- i Después de una prueba o entrenamiento lere el reemplazo del glucógeno muscular. Du- de fondo, es importante consumir una rante las primeras 2 horas después del ejercicio, cantidad sustancial de hidratos de carbo- el ritmo de resíntesis del glucógeno muscular os- no que reemplace el glucógeno consu- cila entre el 7% y el 8% por hora (de 7 a 8 mmol mido durante la actividad. El cuerpo pa- por kg de músculo por hora), lo cual es algo más rece estar preparado para reemplazar el rápido que el ritmo normal de entre el 5% y el glucógeno durante las primeras horas 6% por hora.33 Por lo tanto, un deportista que se después del entrenamiento o competi- recupera de una prueba agotadora que requiere ción, porque los niveles de glucogeno- capacidad de resistencia debe ingerir suficientes sintasa están al máximo. hidratos de carbono tan pronto como sea posible después del ejercicio. También se postuló que añadir proteínas y aminoácidos a los hidratos de carbono ingeridos durante el período de recupera- ción mejoraría la síntesis de glucógeno muscular por encima del nivel conseguido sólo con hidratos de carbono. Varios estudios posteriores no fueron Diseño de bebidas deportivas capaces de confirmarlo,36, 53 pero un estudio más re- Anteriormente, hemos mencionado que ingerir ciente ha demostrado claramente que un suple- soluciones de hidratos de carbono durante el mento de hidratos de carbono y proteínas es más ejercicio puede beneficiar el rendimiento al ase- eficaz para recuperar el glucógeno muscular des- gurar una cantidad adecuada de combustible pa- pués del ejercicio que sólo un suplemento de hi- ra la producción de energía y de fluidos para la dratos de carbono.32 470
- 14. 03 Cap 16-21 • 549-776 17/1/07 16:38 Página 622 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE EE.UU. no tenían un buen papel en la competi- clara desventaja. Pero si los factores neurales son ción internacional debido en gran parte a que tan o incluso más importantes que el tamaño, en- eran más débiles que sus competidores. Lenta- tonces el potencial de ganancia de fuerza absolu- mente, las investigaciones demostraron que las ta es considerable. Asimismo, las mujeres pueden mujeres pueden beneficiarse considerablemente conseguir una hipertrofia muscular significativa. de los programas de entrenamiento de fuerza y Esto se ha demostrado en mujeres culturistas que que las ganancias de fuerza no van acompañadas no han usado esteroides anabólicos. Además, en generalmente de grandes aumentos en el volu- un estudio, hombres y mujeres mostraron aumen- men muscular. tos de un porcentaje similar tanto en la fuerza co- Un estudio comparó las respuestas al entrena- mo en la hipertrofia muscular después de 16 sema- miento de 47 mujeres y 26 hombres que volunta- nas de entrenamiento resistido.15 Los mecanismos riamente participaron en programas idénticos de básicos que permiten mayores niveles de fuerza entrenamiento de pesos con resistencia progresiva. no han sido definidos con claridad, por lo que no El programa se llevó a cabo dos veces por semana, podemos sacar conclusiones en este momento. 40 min al día, durante un total de 10 semanas. Las En general, aunque las mujeres suelen crear sus- ganancias de fuerza fueron las siguientes: tancialmente menos masa muscular que los hom- bres como respuesta a un estímulo dado, algunas • Fuerza en press de banca: 29% en las muje- mujeres adquieren más masa muscular que algu- res y 17% en los hombres. nos hombres. Al igual que la capacidad aeróbica, • Fuerza en press de piernas: 30% en las mu- de la que hablamos previamente, se produce algún jeres y 26% en los hombres. solapamiento entre mujeres y hombres. Teniendo todo esto presente, resulta intere- La circunferencia muscular aumentó sólo li- sante reparar en los récords mundiales masculi- geramente en las mujeres, pero los hombres exhi- nos y femeninos de halterofilia por categorías de bieron una clásica hipertrofia muscular.65 Por lo peso. La figura 18.7 muestra estas plusmarcas tanto, la hipertrofia no es una consecuencia nece- mundiales para el peso total levantado (la suma saria ni un prerrequisito para las ganancias de de la arrancada y los dos tiempos) de 2002. Se fuerza muscular. Diversos estudios han confirma- aprecia en esta figura que los hombres son bas- do estos resultados. tante más fuertes en todas las categorías por el Las mujeres tienen el potencial para desarro- peso. En la parte más baja de la clasificación, los llar sustancialmente más fuerza de la que normal- hombres levantan casi 100 kg más que las muje- mente se identifica en la mujer media, general- res, aunque también levantan 120 kg más en las mente sedentaria. Pero, ¿serán capaces alguna categorías más altas por el peso. Parte de la dife- vez las mujeres de conseguir tener la misma fuer- rencia se explica porque, para un peso corporal za que los hombres para todas las regiones im- dado, es muy probable que los hombres tengan portantes del cuerpo? Tal como hemos tratado más masa magra. Además, pocas mujeres partici- previamente en este capítulo, las proporciones pan en la halterofilia de competición. Cuanto ma- entre fuerza y peso de las piernas entre los sexos yor sea el número de participantes, mayor es la son similares, lo que sugiere que las cualidades posibilidad de conseguir plusmarcas más altas. histológicas y bioquímicas de los músculos son No obstante, las diferencias son tan grandes que las mismas, con independencia del sexo. Esto se deben intervenir otros factores. confirmó mediante tomografías computarizadas de la parte superior de los brazos y de los muslos de estudiantes de educación física de ambos se- xos y de culturistas masculinos.52 Aunque ambos Las mujeres pueden experimentar im- grupos de hombres tenían niveles absolutos de portantes aumentos de fuerza (del 20% fuerza mucho mayores que las mujeres, no se ha- al 40%) como consecuencia del entrena- llaron diferencias entre los grupos cuando la miento resistido, y la magnitud de estos fuerza se expresaba por unidad de área muscular cambios es similar a la observada en los cruzada (figura 18.6). hombres. Probablemente, estas ganan- Debido a sus menores niveles de testostero- cias se deben en mayor medida a factores na, las mujeres tienen menos masa muscular to- neurales, puesto que el incremento de la tal. Si la masa muscular es el principal determi- masa muscular suele ser pequeño. nante de la fuerza, entonces las mujeres están en 622
- 15. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:17 Página XIV CRÉDITOS FOTOGRÁFICOS Figuras 0.0, 0.6a, 0.7a Por cortesía de American Figura 19.10 © 2002 iPhotoNews.com College of Sports Medicine Archives. Reserva- Introducción: Fotografía por cortesía de Cos- dos todos los derechos. med, Srl; © Empics; Fotografía por cortesía de Figura 0.1, 0.2, 0,3a-b, 0.4a-b, 0.5a-c, 0.6b, los autores. 0.7b-c, 0.8a-c, 0.9a-b, 0.12, 1.1a-c, 1.10a-b, Capítulo 1: © Custom Medical Stock Photo; 1.11, 1.12a-c, 1.13a-b, 3.4a-b, 4.20a-b, 6.1, 6.3a- © Human Kinetics; Fotógrafo/ Bruce Coleman, b, 6.6, 10.3a-b, 17.6, 19.1, 19.2, 19.4a-b, 20.7, Inc. 21.9a-c, y fotografías de la contraportada de los autores: Por cortesía de los autores. Capítulo 2: © Custom Medical Stock; © Human Kinetics; © Empics. Figura 0.10, 0.11, 3.2, 4.12, 9.4a-b, 12.11, 12.13, y 19.3 © Tom Roberts. Capítulo 3: © Human Kinetics; © Empics; © Human Kinetics. Figura 1.4, 2.2a, 7.4a, y 20.3a-b © Custom Me- dical Stock Photo. Capítulo 4: © Human Kinetics; © Tom Roberts; © Human Kinetics. Figura 3.1, 3.15, 3.16 a-c, 14.05, 14.06, 18.12, 19.5 y 19.8 © Human Kinetics. Capítulo 5: © Empics; © Human Kinetics; © Empics. Figura 3.10 y 3.11a-b De F.C. Hagerman, R.S. Hikida, R.S. Staron, W.M. Sherman y D.L. Cos- Capítulo 6: © Human Kinetics; © Human Kine- till, 1984. “Muscle damage in marathon runners,” tics; Fotografía por cortesía de los autores. Physician and Sportsmedicine 12:39-48. Capítulo 7: Custom Medical Stock Photo; Figuras 0.8 Fotógrafo/Bruce Coleman, Inc. © Human Kinetics; © Human Kinetics. Figuras 0.9 y 19.9 © Empics. Capítulo 8: Fotógrafo/Bruce Coleman, Inc.; © Human Kinetics; © Human Kinetics. Figura 11.9 Fotografía por cortesía de Dennis Graver. Capítulo 9: © Empics; © Human Kinetics; © Human Kinetics. Figura 11.14. Por cortesía de NASA. Capítulo 10: Fotografía por cortesía de los auto- Figura 14.2a Tom Pantages. res; Fotógrafo/Bruce Coleman, Inc.; © Human Figura 14.3a-b Fotografía por cortesía de Holo- Kinetics. gic, Inc. Capítulo 11: © Empics; © Digital Vision; Corte- Figura 14.4. Fotografía por cortesía de Life Me- sía de NASA. asurement, Inc. Capítulo 12: Fotógrafo/Bruce Coleman, Inc.; Figura 18.10a-b Reproducido de J. Bone Miner © Empics; © Human Kinetics. Res 1986, 1:15-21 con autorización de la Ameri- Capítulo 13: © Human Kinetics; © Human Ki- can Society for Bone and Mineral Research. Fo- netics; © Human Kinetics. tografías por cortesía de D.W. Dempster. Capítulo 14: Tom Pantages; © Human Kinetics; © Human Kinetics. XIV
- 16. 03 Cap 16-21 • 549-776 17/1/07 16:38 Página 575 NIÑOS Y ADOLESCENTES EN EL DEPORTE Y EN EL EJERCICIO 12. Inbar, O., & Bar-Or, O. (1986). Anaero- Bibliografía bic characteristics in male children and adoles- cents. Medicine and Science in Sports and Exer- 1. Bar-Or, O. (1983). Pediatric sports medi- cise, 18, 264-269. cine for the practitioner: From physiologic prin- 13. Kraemer, W.J., & Fleck, S.J. (1993). ciples to clinical applications. Nueva York: Strength training for young athletes. Champaign, Springer-Verlag. IL: Human Kinetics. 2. Bar-Or, O. (1989). Temperature regulation 14. Krahenbuhl, G.S., Morgan, D.W., & during exercise in children and adolescents. En Pangrazi, R.P. (1989). Longitudinal changes in C.V. Gisolfi & D.R. Lamb (eds.), Perspectives in distance-running performance of young males. exercise science and sports medicine: Youth, International Journal of Sports Medicine, 10, 92- exercise and sport (págs. 335-362). Carmel, IN: 96. Benchmark Press. 15. Mahon, A.D., & Vaccaro, P. (1989). Ven- 3. Bass, S.L. (2000). The prepubertal years: tilatory threshold and nVO2max changes in chil- A uniquely opportune stage of growth when the dren following endurance training. Medicine and skeleton is most responsive to exercise? Sports Science in Sports and Exercise, 21, 425-431. Medicine, 30, 73-78. 16. Malina, R.M. (1989). Growth and matu- 4. Bjorntorp, P. (1986). Fat cells and obesity. ration: Normal variation and effect of training. En K.D. Brownell & J.P. Foreyt (eds.), Handbo- En C.V. Gisolfi & D.R. Lamb (eds.), Perspecti- ok of eating disorders: Physiology, psychology, ves in exercise science and sports medicine: and treatment of obesity, anorexia, and bulimia Youth, exercise and sport (págs. 223-265). Car- (págs. 88-98). Nueva York: Basic Books. mel, IN: Benchmark Press. 5. Boisseau, N., & Delamarche, P. (2000). 17. Malina, R.M., & Bouchard, C. (1991). Metabolic and hormonal responses to exercise in Growth, maturation, and physical activity. children and adolescents. Sports Medicine, 30, Champaign, IL: Human Kinetics. 405-422. 18. Pfeiffer, R.D., & Francis, R.S. (1986). 6. Brooks, G.A., Fahey, T.D., White, T.P. y Effects of strength training on muscle develop- Baldwin, K.M. (2000). Exercise physiology: Hu- ment in prepubescent, pubescent, and postpubes- man bioenergetics and its applications, 3ª ed. cent males. Physician and Sportsmedicine, 14(9), Mountain View, CA: Mayfield. 134-143. 7. Clarke, H.H. (1971). Physical and motor 19. Ramsay, J.A., Blimkie, C.J.R., Smith, tests in the Medford Boys’ Growth Study. Engle- K., Garner, S., MacDougall, J.D., & Sale, D.G. wood Cliffs, NJ: Prentice-Hall. (1990). Strength training effects in prepubescent 8. Daniels, J., Oldridge, N., Nagle, E, & boys. Medicine and Science in Sports and Exer- White, B. (1978). Differences and changes in cise, 22, 605-614. nVO2 among young runners 10 to 18 years of age. 20. Robinson, S. (1938). Experimental stu- Medicine and Science in Sports and Exercise, 10, dies of physical fitness in relation to age. Ar- 200-203. beitsphysiologie, 10, 251-323. 9. Eriksson, B.O. (1972). Physical training, 21. Rogers, D.M., Olson, B.L., & Wilmore, oxygen supply and muscle metabolism in 11-13- J.H. (1995). Scaling for the nVO2-to-body size re- year old boys. Acta Physiologica Scandinavica, lationship among children and adults. Journal of (Supl. 384) 1-48. Applied Physiology, 79, 958-967. 10. Fleck, S.J., & Kraemer, W.J. (1987). De- 22. Rowland, T.W. (1985). Aerobic response signing resistance training programs 2ª ed. to endurance training in prepubescent children: A Champaign, IL: Human Kinetics. critical analysis. Medicine and Science in Sports 11. Froberg, K., & Lammert, O. (1996). De- and Exercise, 17, 493-497. velopment of muscle strength during childhood. 23. Rowland, T.W. (1989). Oxygen uptake En O. Bar-Or (ed.), The child and adolescent ath- and endurance fitness in children: A developmen- lete (pág. 28). Londres: Blackwell Science. 575
- 17. 01 Crédits+Cap 01-06 • 001-220 17/1/07 16:19 Página 88 FISIOLOGÍA DEL ESFUERZO Y DEL DEPORTE 3. Marieb, E.N. (1995). Human anatomy Enoka, R.M., & Stuart, D.G. (1984). Henne- and physiology (3.ª ed.). Nueva York: Benja- man’s “size principle”: Current issues. Trends in min/Cummings. Neurosciences, 7, 226-227. 4. Petajan, J.H., Gappmaier, E., White, A.T., Gandevia, S.C. (1996). Kinesthesia: Roles for Spencer, M.K., Mino, L., & Hicks, R.W. (1996). afferent signals and motor commands. En L.B. Impact of aerobic training on fitness and quality Rowell & J.T. Shepherd (Eds.), Handbook of of life in multiple sclerosis. Annals of Neurology, physiology: Section 12. Exercise: Regulation and 39, 432-441. integration of multiple systems (págs. 128-172). 5. Pette, D., & Vrbova, G. (1985). Neural Nueva York: Oxford University Press. control of phenotypic expression in mammalian Gielen, C.C.A.M., & Denier van der Gon, J.J. muscle fibers. Muscle and Nerve, 8, 676-689. (1990). The activation of motor units in coordi- nated arm movements in humans. News in Phy- siological Sciences, 5, 159-163. Hasan, Z., Enoka, R.M., & Stuart, D.G. (1985). The interface between biomechanics and Lecturas seleccionadas neurophysiology in the study of movement: So- me recent approaches. Exercise and Sport Scien- Bawa, P. (2002). Neural control of motor out- ces Reviews, 13, 169-234. put: Can training change it? Exercise and Sport Henneman, E., & Mendell, L.M. (1981). Sciences Reviews, 30, 59-63. Functional organization of motoneuron pool and Binder, M.D., Heckman, C.J., & Powers, its inputs. En V.B. Brooks (ed.), Handbook of R.K. (1996). The physiological control of moto- physiology. Section I, Volume II, The nervous sys- neuron activity. En L.B. Rowell & J.T. Shepherd tem: Motor control, Part I (págs. 423-507). Bet- (Eds.), Handbook of physiology: Section 12. hesda, MD: American Physiological Society. Exercise: Regulation and integration of multiple McArdle, W.D., Katch, F.I., & Katch, V.L. systems (págs. 3-53). Nueva York: Oxford Uni- (2001). Exercise physiology: Energy, nutrition, versity Press. and human performance (5ª ed.). Baltimore: Wi- Christensen, N.J., & Galbo, H. (1983). Sym- lliams & Wilkins. pathetic nervous activity during exercise. Annual O’Donovan, M.J. (1985). Developmental regu- Review of Physiology, 45, 139-153. lation of motor function: An uncharted sea. Medici- Cope, T.C., & Pinter, M.J. (1995). The size ne and Science in Sports and Exercise, 17, 35-43. principle: Still working after all these years. Powers, S.K., & Howley, E.T. (2001). Exerci- News in Physiological Sciences; 10, 280-286. se physiology: Theory and application to fitness Edgerton, V.R., Bodine-Fowler, S., Roy, R.R., and performance (4ª ed.). Madison, WI: Mc- Ishihara, A., & Hodgson, J.A. (1996). Neuromus- Graw-Hill. cular adaptation. En L.B. Rowell & J.T. Prochazka, A. (1996). Proprioceptive feed- Shepherd (Eds.), Handbook of physiology: Sec- back and movement regulation. En L.B. Rowell tion 12. Exercise: Regulation and integration of & J.T. Shepherd (Eds.), Handbook of physiology: multiple systems (págs. 54-88). Nueva York: Ox- Section 12. Exercise: Regulation and integration ford University Press. of multiple systems (págs. 89-127). Nueva York: Emonet-Denand, F., Hunt, C.C., & Laporte, Y. Oxford University Press. (1988). How muscle spindles signal changes of Proske, U. (1997). The mammalian muscle muscle length. News in Physiological Sciences, spindle. News in Physiological Sciences; 12, 37- 3, 105-109. 42. Enoka, R.M. (2001). Neuromechanics of hu- Roy, R.R., Baldwin, K.M., & Edgerton, V.R. man movement (3ª ed.). Champaign, IL: Human (1991). The plasticity of skeletal muscle: Effects Kinetics. of neuromuscular activity. Exercise and Sport Sciences Reviews, 19, 269-312. 88