
Recomendados
Recomendados
Mais conteúdo relacionado
Mais procurados
Mais procurados (20)
Semelhante a Tema 6 Modificaciones postraduccionales
Semelhante a Tema 6 Modificaciones postraduccionales (20)
Mais de juanvelez22
Último
Último (20)
Tema 6 Modificaciones postraduccionales
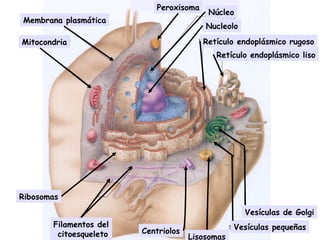
- 1. Nucleolo Retículo endoplásmico rugoso Retículo endoplásmico liso Vesículas de Golgi Vesículas pequeñas Membrana plasmática Mitocondria Centriolos Ribosomas Filamentos del citoesqueleto . . . . . . . . . . . . . Peroxisoma Núcleo . . . . Lisosomas .
- 8. Proteína Disulfuro Isomerasa PDI
- 16. 13 – 36 residuos hidrofóbicos
- 18. Type I: Signal sequence on amino terminus enters first and continues to elongate. Protein is threaded through the translocating channel (open area in rer membrane) until a hydrophobic stop sequence is reached. That hydrophobic stop sequence (seen as a hatched region in the protein) is then inserted in the membrane and forms the anchor for that protein. Signal is cleaved by protease inside the lumen.
- 19. Type II: No cleavable signal sequence. These proteins have rather long hydrophobic regions that will be anchored in the membrane. Type II proteins are threaded into the lumen with the C terminus leading. Protein continues to be inserted until it reaches the hydrophobic stop signal sequence. Type III: Same as Type II, only the N terminus leads into the lumen.
- 20. The "positive inside rule" states that amino acid residues nearest the cytosolic side of the hydrophobic anchor sequence are more positive than those nearest the lumenal side. So, whichever end has the least positive charges near the signal anchor patch would go into the ER lumen. One can change the direction of translocation of a protein (reverse it) by mutating the protein and making more positively charged groups near the anchor patch of the other end. Below, the cartoon shows that this can be done to change a Type II protein (COOH end enters ER lumen) to a Type III (which has its amino terminal entering the lumen).
- 21. Washburn and Speiss (JCB 137: 555-562, 1997) also tested the length of the hydrophobic signal anchor sequence. The following cartoon shows that a longer hydrophobic anchor sequence (seen as the portion running through the membrane) promotes entry with the amino terminal leading into the lumen.
- 24. N – N-Acetilglucosamina
- 25. S –T (OH) N-Acetil-galactosamina OH-K Galactosa
- 33. Cis-Golgi
- 35. The congenital disorders of glycosilation (CDG) were originally called carbohydrate-deficient glycoprotein syndromes (CDGS) and are a subset of genetic defects affecting primarily N-glycan assembly.
- 40. Catepsina D y otras enzimas lisozomales
- 49. RER 3 min Aparatp de Golgi 20 min Vesículas secretoras 90 min Liberación 120 min Páncreas tritio* Radio-autografía
- 51. RER Cis - Golgi Fijación grupo fosfato a manosas en enzimas lisosómicas evita que sufran nuevos cambios Remueve manosas de glicoproteínas . Fija manosas N-oligosacáridos (N) Red Cis Vesículas de transición
- 52. Medial - Golgi Remueve manosas por manosidasas I y II adiciona N-acetilglucosamina por N-acetil-glucosamina transferasa I y II
- 53. Trans - Golgi Completa la gricosilación, adición de galactosa y ácido siálico (transferasas) O-oligosacaridos (S/T/HO-K) Liberación de proteínas Red Trans Vesículas de transición .
- 60. Acetilacion
- 61. The chemical conversion of arginine to citrulline, known as citrullination or deimination. These citrulline residues are generated by a family of enzymes called peptidylarginine deiminases (PADs) , which convert arginine into citrulline in a process called citrulination or deimination .
- 64. Certain proteins are anchored to biological membranes by lipid anchors. Particularly common are the N-myristoyl – and S-palmitoyl – anchoring motifs shown here . N-Myristoylation always occurs at an N-terminal glycine residue , whereas thioester linkages occur at cysteine residues within the polypeptide chain. G-protein – coupled receptors, with seven transmembrane segments, may contain one (and sometimes two) palmitoyl anchors in thioester linkage to cysteine residues in the C-terminal segment of the protein.
- 65. Some proteins have lipid moieties attached: The viral src protein is myristoylated at the N-terminal glycine. Rhodoposin is palmitoylated at a cysteine residue The ras oncogene protein is farnesylated as are some G proteins. Some eukaryotes, notably parasitic protozoa, have glycosylphosphatidylinositol -linked proteins
- 66. Proteins containing the C-terminal sequence CAAX can undergo prenylation reactions that place thioether-linked farnesyl or geranylgeranyl groups at the cysteine side chain. Prenylation is accompanied by removal of the AAX peptide and methylation of the carboxyl group of the cysteine residue, which has become the C-terminal residue.
- 67. The glycosyl phosphatidylinositol (GPI) moiety is an elaborate lipid-anchoring group. Note the core of three mannose residues and a glucosamine. Additional modifications may include fatty acids at the inositol and glycerol OOH groups.
- 69. A two dimensional representation of the Human Aquaporin 0 (AQP0) structure indicating sites of truncation (arrows), sites of deamidation (green residues) and sites of phosphorylation (red diamonds).
- 70. Chymotrypsin and trypsin are both synthesized as zymogens . Cleavage of chymotrypsinogen between Arg15 and Ile 16 by trypsin yields the enzymatically active pi-chymotrypsin. Two further proteolytic cleavages catalyzed by chymotrypsin removes the dipeptides Ser14-Arg15 and Thr147-Asn148 to yield alpha-chymotrypsin. Trypsin is activated by the removal of the N-terminal seven amino acids.
- 71. Proteins that require a prosthetic group for activity must have this group added. For example, the haem (heme) group must be added to globins and cytochromes; Fe-S clusters must be added to ferredoxins.
- 77. Antes Después Antes Después